
Use the Search magnifying glass above to search the site. Use CNTL+F to find text on pages; Apple, use Command+F.
COHERENCE: The Human Body & Our Electrical Relationship With Planet Earth

COHERENCE, Vol 1, Issue 4 , January, 2023, by Stephen Elliott, Life Scientist, COHERENCE
Keywords: Bio-current, breathing, cellular respiration, Clinton Ober, coherence, Coherent Breathing®, current, Earth, earthing, EEG, electron transport chain, grounding, human body, mitochondria, Nabhi Kriya, planet Earth, respiration, respiratory chain, Stephen Elliott, voltage.
Undergoing evolution. Last revised on September 2nd, 2023.
Preface: This article explores the apparent attraction in connectivity between the human body and Earth ground. Exploration of this topic by this author began in 2004, resulting in the filing of US Patent 7349194 in 2005, Method And System For Electrically Connecting The Human Organism (And Vertebrate) Organism To Earth So As To Facilitate A Current Flow Between The Human Biopotential And Earth For The Purpose Of Promoting Health, Well Being, and Performance.
The research behind the patent application demonstrated that the human body desires connectivity with Earth between a range of resistances, where current flow is maximized when resistance between the body and Earth is less than 500 Ohms, but is negated rapidly when resistance values rise above this value. Further investigation of the meaning and significance of the phenomenon was not undertaken until the writing of this article, which began in 4Q2022.
The present hypothesis regarding the phenomenon being observed is that we are measuring electron flow from Earth to the body, and that this electron flow is a consequence of metabolic status, where these electrons are being conscripted by the electron transport chain of the mitochondria associated with “cellular respiration”.
For those familiar with my writing of the last 17 years, you will know that I have been interested in the inner vs. outer workings of “breathing”, diving ever deeper into circulatory, gravitational, and energetic aspects. While the present line of inquiry is not about breathing per se, it does appear that it intersects with the quest to understand breathing, now stepping inside vs. outside the cell proper. Who could know that these disparate pursuits would intersect in the cell? Not I.
There are a couple of significant matters that arise, right or wrong yet to be determined: 1) It is generally considered that the human body receives all of the electrons it requires for cellular respiration via dietary and respiratory intake. If this is true, why does the body appear to want to maximize the flow of electrons from Earth given the chance? Here, my hypothesis is that electrons external to the body with <500 ohms reach require less work to recruit than electrons available internally.
A 2nd major question is, if when the body is connected electrically to a ground rod, we have hundreds of trillions of electrons per second flowing from Earth into the body in what is primarily a DC current, there must exist a circuit by which electrons return to Earth. Where is it? The only obvious answer is that we expel electrons into the atmosphere with each exhalation of breath, which would necessitate that we also inhale electrons with each breath, which is already assumed.
~
As introduction, I am a proponent of “grounding”. I sleep grounded, and at work in my home office I am grounded. I have been interested in this topic for much of my life, it always making sense to me that before modern times, we were all grounded, during sleep, rest, activity. This because we were sleeping, walking, sitting on the ground, the Earth’s surface. Whereas today, our modern lifestyles have us electrically insulated from Earth, via the materials of our shoes, our clothing, our homes, our furniture, and our activities, where human agriculture, working the Earth with our hands and bodies used to be necessary to sustain our lives. So, we were in constant contact with the Earth’s surface, be it soil, water, plant life…
It is generally understood that the human race populated the planet by walking the ocean’s shorelines, from central Africa, to the shorelines of the farthest reaches of the world, parts of the planet that are now separated by ocean, e.g. the Bering Straight, the waters separating Australia from mainland Asia. Humankind reached every corner of the world by this endless quest. The import of this is that people were moving and living next to the sea, a highly conductive saline environment, damp rock, sand, earth, water, all highly conductive relative to humankind’s electrical connectivity with the Earth. The sea is in effect “liquid ground”.
By and large, this was the best place to be because both food and fresh water were plentiful, freshwater streams and rivers emptying into the ocean, fish and predator swimming upstream to capture their prey.
A question that has been in the back of my mind for many years is whether we need connectivity with Earth in order to “function naturally”? Wild animals live outside, exposed to the elements, either standing on Earth, dwelling in the branches, or residing in the water, somehow managing to survive harsh conditions, hot or cold, wet or dry. This includes humankind’s ancient ancestors that have had an evolutionary trajectory of our own. How do they do it?
The question is: “Do we as “civilized” humans require connectedness to Earth?” Does the human organism desire it, from a homeostatic point of view? Of course, a larger question is: “Is there communication going on between people and the planet?” (The same question of other life forms…?)
Clinton Ober’s book: Earthing: The Most Important Health Discovery Ever!, copyright 2014 (1), considers many of the same ideas. However, I am not aware of anyone employing this approach to understanding it.
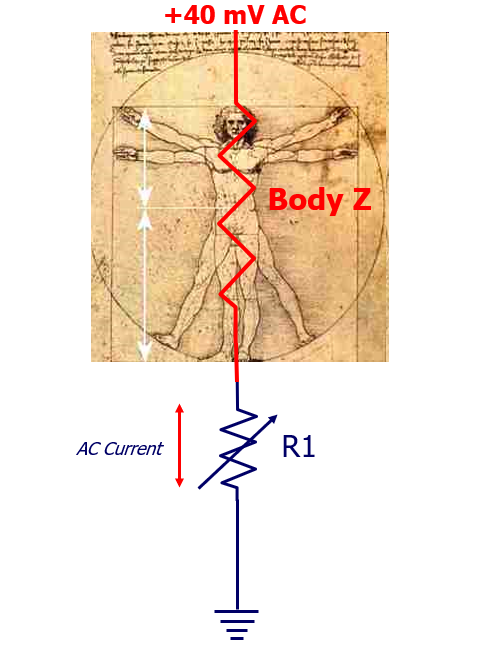
Figure 1: The Measurement Concept
To explore this question, I set about to see how the human body responds to “connectivity” with ground”. To start, I measured the AC voltage produced by the human body, which measures ~+40mVAC; +.040 volts, “alternating current” to ground. (The instrument, an Agilent 3458A, has a 1M Ohm input impedance so it is capable of measuring very small signals without loading them down, i.e., without consequent change in value.) Note that the potential is positive relative to ground.
My objective here is to assess “AC current”, but because the signals are very small, I could not place a current meter directly in-line with the human body and realize any meaningful readings. On the other hand, because the instrument I am using is capable of measuring very small voltages, I am able to assess AC voltage across resistor R1, a resistor decade box, and from there use Ohm’s Law to determine AC current. I am specifically interested in “AC” activity because I am fascinated with the potential for communication.
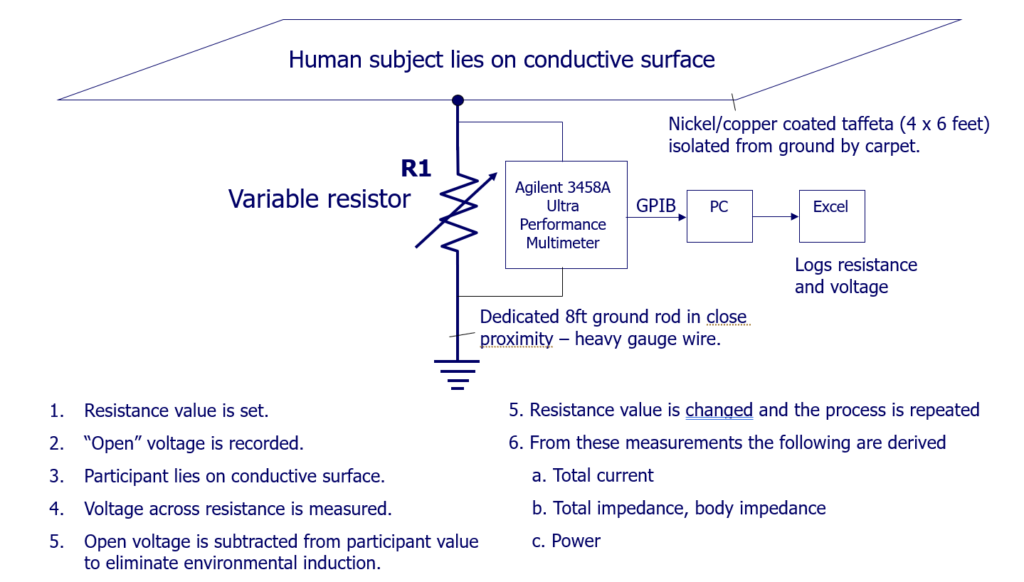
Figure 2: The Measurement System
The measurement system is depicted in Figure 2, where two measurements are required per resistance value, the first set of measurements taken without a person lying on the conductive surface, a 4 x 6 piece of copper/nickel coated taffeta, insulated from ground via carpet. The second set of measurements are taken with a human subject lying on the surface, without clothing, i.e., maximizing contact between the body and the fabric.
The first set of measurements are then subtracted from the second, the intention being to eliminate any environmentally induced “noise”.
One can see that having the “source” voltage of a human subject, a known value of resistor R1, and the calculated current flowing through R1, we are able to know if human body impedance is changing as a consequence of the resistor value, i.e. “connectedness” to Earth. This is tantamount to asking: Does the human body have a characteristic impedance? If so, what is it?
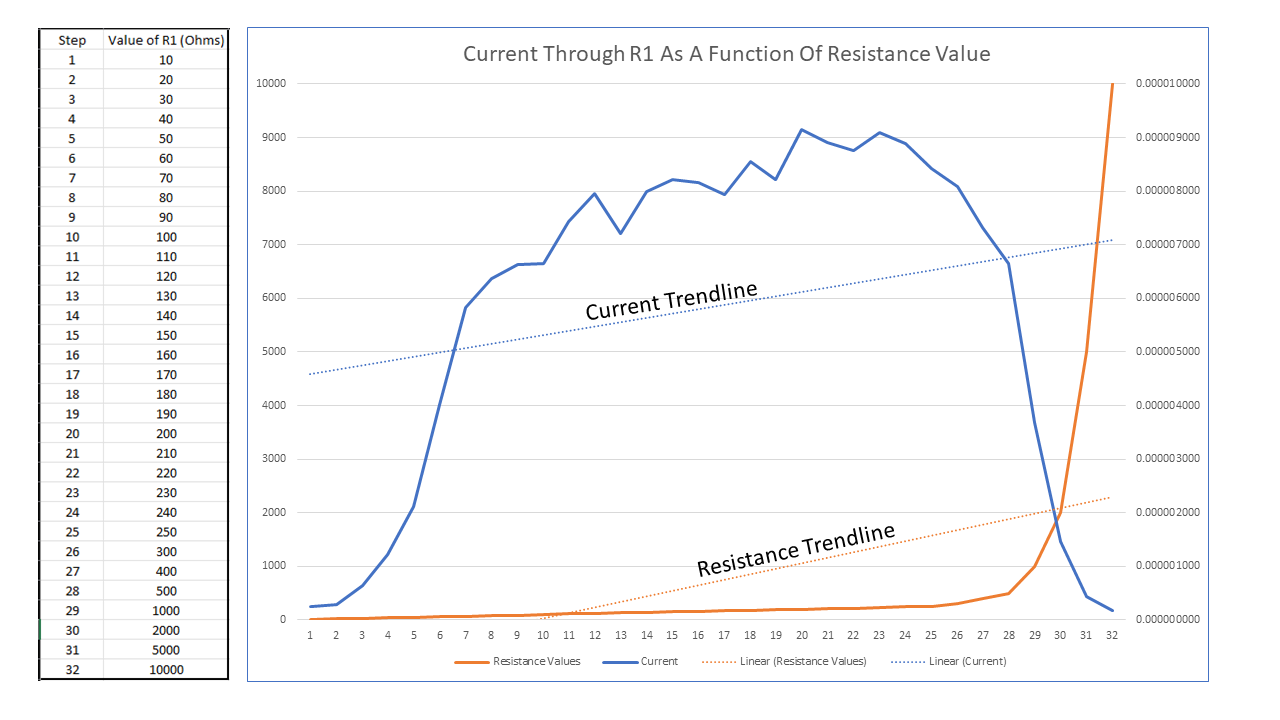
Figure 3: Current Through R1 As Function Of Resistance Value
Figure 3 presents what is in my view a 1st seminal test regarding this inquiry. Here we have resistance values increasing in 32 steps from 10 ohms up to 10,000 ohms (left Y axis, X axis), the orange curve, and their consequent AC current values, (right Y axis), the blue curve.
Here we see current rising rapidly as we progress from 10 ohms up to 70 ohms, after which it begins following an upward linear progression with current increasing as resistance increases up to 200 ohms, after which it begins a rapid decline, current approximating zero amps at 10,000 ohms. Here, I posit that measurements south of 100 ohms are not a legitimate reflection of current, but that the value is being “shunted” to ground, i.e., overloaded due to the low resistance values.
However, I do believe values between 100 ohms and 200 ohms to be a true reflection of the body’s response to “connectivity” to Earth ground. Note that the linear trendlines of Current and Resistance are parallel, i.e., correlative.
(Here please note that I am using a dedicated ground rod for these measurements, i.e. it is not bonded in any way to the electrical grid. This and I advise against use of the ground that is bonded to the electrical grid, as is required by electrical code for safety. Secondly, be cautious regarding the condition and proximity of electrical devices, e.g. a bedside lamp. Be sure that they are in safe electrical condition and that there is no opportunity for shock hazard.)
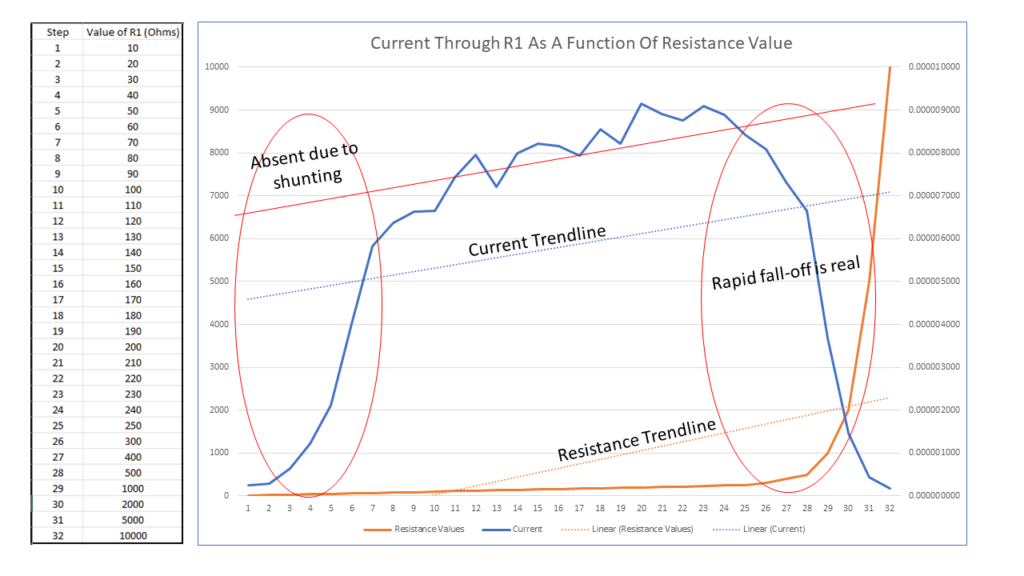
Figure 4: Interpretation
Figure 4 makes clear my interpretation of these results.
If the current offered by the body remains relatively constant as resistance increases, then we can expect the voltage across R1 to increase as resistance values increase, i.e., for the same current, the voltage across a higher value resistor will be higher. This appears to be what is happening…
The fact that current is increasing as resistance is increasing between 70 and 200 ohms is especially significant. If the electrical potential and impedance of the body were constant, like a battery, this would not be happening. Instead, current would be decreasing as resistance is increasing. Instead current appears to be increasing in this 70-200 ohm range. I posit that this is an “active” biological response in order to maximize connectivity, i.e., maximize current flow with Earth.
So, I presently consider the solid red line to be a sound estimate of the persistent current that the body is offering, even as resistance is increasing – until the precipitous drop above 200 ohms. This relatively constant AC current value is on the order of 8.5 micro-amps. This makes sense, but what of the precipitous fall above 200 ohms? Why is this happening?
To understand, we need to widen our scope to bring “impedance” into view.
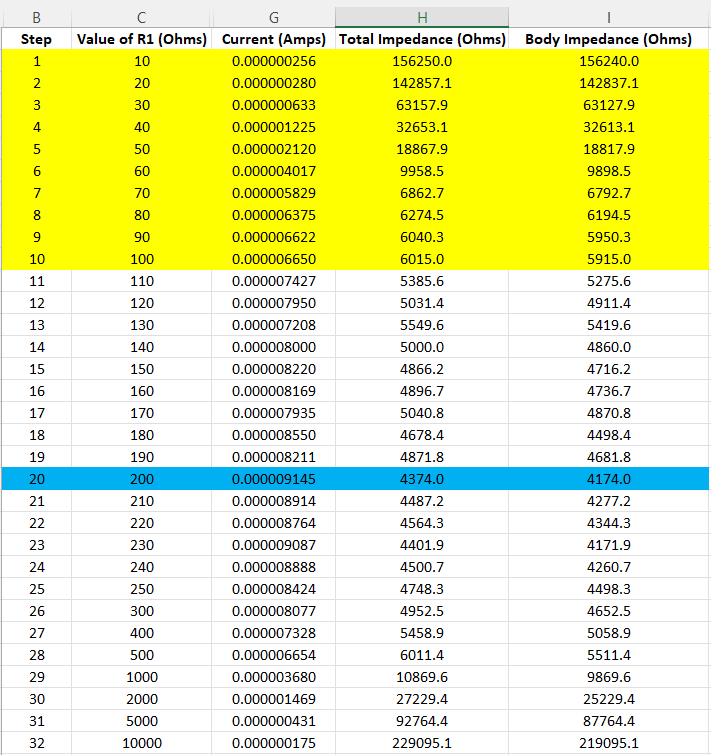
Figure 5: Resistance, Current, & Impedance
Figure 5 presents the columns H and I, “Total Impedance” and “Body Impedance”. Total impedance is based on the initial determination that the “source” body voltage is 40mVAC. Therefore, if we divide this source voltage by the current that is assessed at each resistance value, we arrive at “Total Impedance” at that value, column H.
If we subtract the value of the resistor we arrive at Column I, “Body Impedance”. If we discount the area below 100 ohms (yellow), because it is an outcome of shunting the signal (current hypothesis), and plot values of columns C and I above 100 ohms, it yields the graphs of Figure 6.
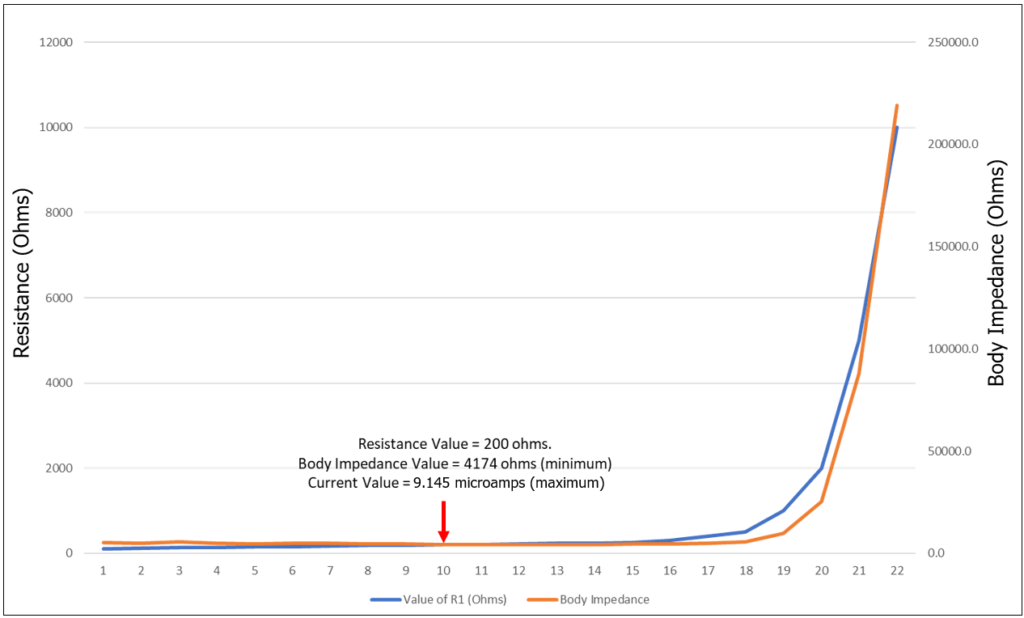
Figure 6: Resistance vs. Body Impedance
Figure 6 makes the correlation very clear. Body impedance bottoms out at 200 ohms and AC current is maximized at 9.145 microamps (see row 20 of table of Figure 5). The impedance of the body is 4174 ohms. Conversely, as the resistance value passes 500 Ohms, we begin to see a sharp increase in body impedance, from 5511 Ohms at 500, to 40 times that ― 219K Ohms at 10,000.
This result says, that if we connect the human body to a dedicated ground rod, via 200 ohms, current between the body and ground will be maximized. The question remains, to what purpose?
I initially performed this experimentation in 2003-2004, and filed US Patent 7,349,194 in July 2005, which was awarded in March of 2008. Note that sitting grounded was part of my personal meditation protocol at that time, and still is. This was at the height of my pursuit that would come to be trademarked as “Coherent Breathing”.
I repeated the experiment just prior to the writing of this article in January 2023, knowing that my original experiment was flawed due to my use of the electrical “ground” of my home.
In the 2006 timeframe, I drove a dedicated ground rod just outside my office wall and found a substantial difference (261 Ohms of difference relative to the values asserted in the patent application). I credit this difference with “noise” that existed (or exists) on the “power grid”, where noise on the grid is higher in amplitude than the biological signals we are trying to measure. I could see it with an oscilloscope. With the dedicated ground rod, I arrived at 190 Ohms. I presented the findings at the 2009 INSR meeting in Indianapolis with the results that the “resonant” resistance value was 190 Ohms.
My most recent determination is that the resistance that yields the highest current between the human body and Earth is 200 Ohms, very close to the 190 ohms previously asserted.
The key take-away here is that the body appears very responsive to “connectedness to Earth” at values less than 500 Ohms and increasingly non-responsive to values above 500 Ohms. This suggests to me that there is a biological function in play.
~
In 2012, a significant paper considering the health effects of “earthing” was published in The Journal of Environmental and Public Health: Health Implications of Reconnecting the Human Body to the Earth’s Surface Electrons. It is a review of research and papers published by others. Some of the most significant findings are that sleeping grounded for 6 weeks yielded an overall reduction in cortisol levels over a 24 hour period, cortisol being an indication of sympathetic nervous system emphasis. Another study cited, posits that grounding shields the human body from electromagnetic fields induced by 60 Hertz (US standard) radiated by the common electrical wiring of the home or business, a 70% reduction in the 60 Hertz component present in the body.
Most interesting to me is the immediacy of effect reported on autonomic state, where upon grounding there is a prompt shift toward parasympathetic emphasis. This is the reason I was employing it during meditation, i.e. to help bring on autonomic balance and the elicitation of parasympathetic emphasis necessary to meditation. My curiosity led me on to find out if there was a “resonant” characteristic impedance that yielded the optimal exchange of current with ground. This turns out to be 200 ohms. When one is connected this way, one can feel increased current in the body: bioenergy, “qi”. When one is breathing “coherently” an increase in current between body and ground can be observed.
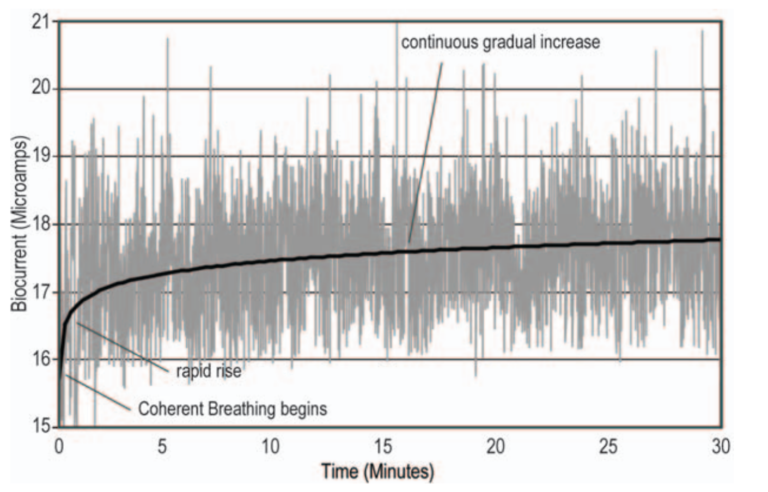
Figure 7: Rapid Rise In Bio-current While Breathing Coherently (Source: The New Science Of Breath, 2005)
Figure 7 presents the rise in bio-current as a consequence of Coherent Breathing. Notice that the rise is very rapid when Coherent Breathing begins, but a gradual rise continues for the duration of measurement.
Why is current rising? Given that Earth ground is our solid reference, it has to be true that Coherent Breathing is resulting in the body becoming more positive as breathing continues, i.e., the bioenergetic potential of the body is increasing where it is the cells of the body, estimated by Medical Physiology (N) as 100 trillion in number, that are generating this potential. (This is certain to be a factor in why resonant breathing is healthful ― more blood flow, more fluid exchange, more oxygenization ― more energy generation…)
Or, is it that the impedance of the body that is decreasing as Coherent Breathing continues, thereby resulting in an increase in current? As we are measuring via the skin, which possesses the “galvanic skin response”, it isn’t the skin that is becoming more conductive. We know this because of previous experiments where it is demonstrated that the skin becomes less conductive from point to point, for example from finger to finger, as one breathes more slowly, deeply, and rhythmically.
This is true of galvanic skin response relative to autonomic status, where Coherent Breathing elicits a parasympathetic state and consequent decrease in skin conductivity (more resistance or less conductivity), where sympathetic response elicits the opposite (less resistance or more conductivity). It is possible that the path from skin to the internal workings of the body has a different response, but we can’t know this without instrumenting the body invasively, an experiment yet to be performed by this author.
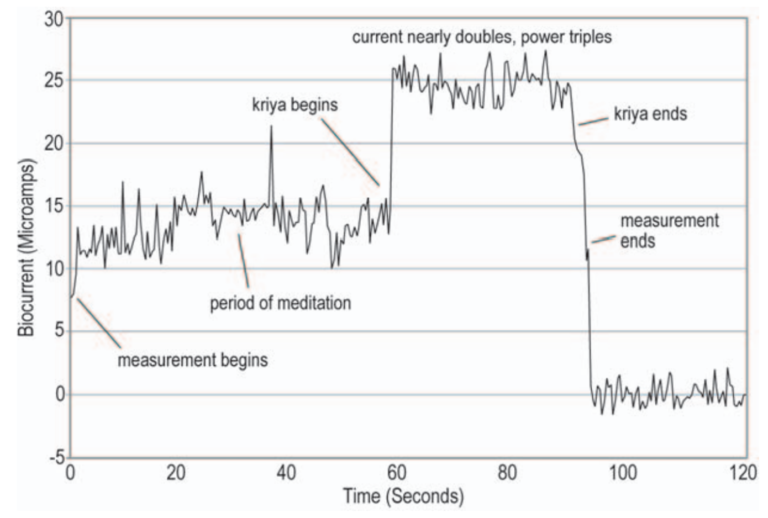
Figure 8: Bio-current Of Spontaneous Kriya (Source: The New Science Of Breath, 2005)
Figure 8 above, demonstrates what can happen when: a) grounded, b) when breathing “coherently”, c) when meditating. Current is rising gradually until, snap, it doubles instantly. When this happens, the body begins to “shake” and “move of its own accord”, hence “spontaneously”. Note that this recording was taken while lying flat, so the movement is that of the spine and limbs undulating in X, Y, and Z directions. If we let it proceed of its own accord without interfering, it will make its way throughout the body, seemingly addressing every aspect of the body where tension resides. Eventually, it will end and the body will be completely relaxed and still.
My initial personal discovery of this occurred while I was grounded, but after learning of it, I determined that it is fundamentally a function of resonant breathing and deep relaxation, i.e. grounding is not necessary. However, as I am now aware of the electrical relationship between the body and Earth, I prefer to do it grounded in order to let the body and Earth have their electrical dialog.
Note that “Coherent Breathing” is a special form of resonant breathing that errs on the parasympathetic side of the autonomic equation. If we accept that .1 Hertz is the nominal frequency of resonance of the average adult, this translating to 6 breaths per minute. Coherent Breathing proper, operates at .085 Hertz, translating to 5.1 breaths per minute. For this reason, it can automatically result in meditation, and with deep conscious relaxation, this “spontaneous kriya”, a cherished Yogic phenomenon associated with shaktipat (divine transmission) and the awakening of kundalini. (4)
~
There is an emerging understanding, though not new, that many diseases, both mental and physical are related to metabolic dysfunction. With some significant exceptions, metabolic rate is a primary determinant of life span, the faster the rate the shorter the lifespan and the slower the rate the longer the lifespan.
Aging is ultimately the accumulation of wear and tear on the body’s metabolic apparatus, and the eventual decline and failure of mitochondria. Mitochondrial failure eventually leads to cell failure and death, “apoptosis”, cell death winnowing away the mass of the body, organs shrinking, height and weight diminishing until death.
Mitochondria are biological nano-engines inside cells that produce energy in the form of ATP (adenosine triphosphate). (6) Mitochondria, estimated to be billions of years old, are thought to pre-date all eukaryotic life forms (life forms composed of cells with a nucleus), this including all forms of life that we can see with the naked eye, plant and animal, due to the fact that eukaryotic life forms could not have existed without mitochondria. This is because mitochondria are responsible for metabolism, on which all forms of eukaryotic life itself depends.
Mitochondria might be thought of as the primordial keepers of the eukaryotic cell as they are involved-in, if not the masters-of many cellular functions including thermal regulation. Interestingly, the distinction between cold-blooded and warm-blooded animals is their metabolic rate, and with their metabolic rate, the number of mitochondria that their cells possess. More energy production results in increased thermal energy produced. (7).
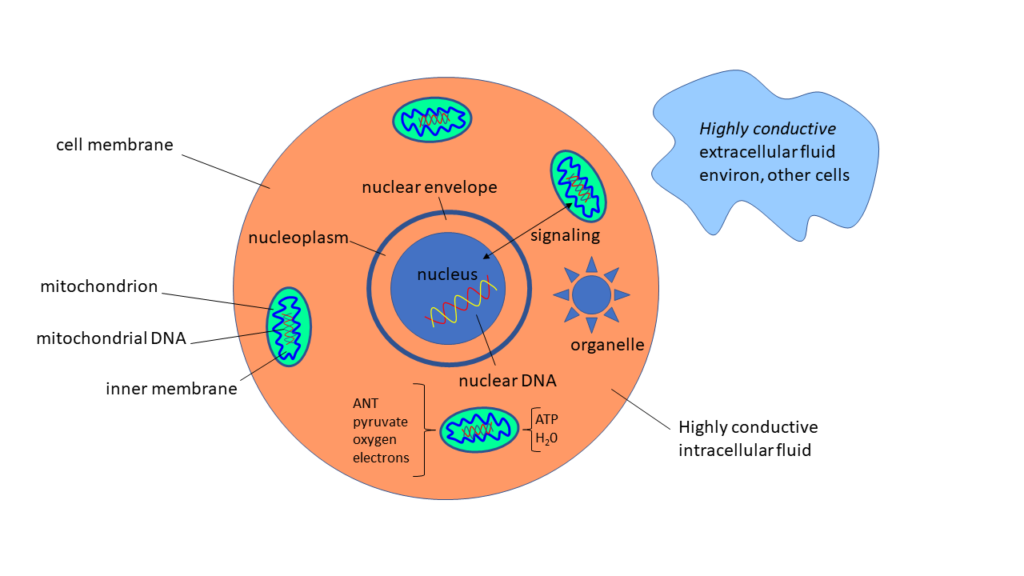
Figure 9: Simplified eukaryotic cell and its attendant mitochondria floating in highly conductive saline environ
The body is estimated to have 100 trillion cells (N), each cell having ~100 mitochondria (N), and each mitochondria having tens of 1000s (N) of unique “respiratory chains” wherein ATP is produced. Primary mitochondrial inputs are pyruvate (produced via glycolysis), oxygen (obtained via breathing), hydrogen (obtained via pyruvate), electrons (obtained via breathing, nutrition, and connectedness with Earth.) Outputs are ATP and water. Note that cells are floating in a highly conductive saline extracellular environment, an environment that facilitates the flow of electrical current.
The combined energy output of the human body can be significant, enough to generate .25 HP or more of work for a sustained period.
.25 HP may not sound like much, but it has been posited that mammalian body, mass per mass, generates more energy than the sun. (N)
The reason why work can be sustained is that energy output can be sustained; the reason why energy output can be sustained is that mitochondria are busy recycling the ATP (adenosine triphosphate) that is used to perform work, ATP->ADP (adenosine diphosphate) and ADP->ATP. The ADP input mechanism is a protein known as (ANT) adenine nucleotide translocator, ANT flowing into the mitochondria as ATP flows out. (N)
Mitochondria perform this task non-stop, lest our lives come to an end. It has also been posited that every day we produce an amount of ATP that approximates our own weight – a mind blowing matter. (8)
Interestingly, many of the health problems that can be attributed to metabolism, and potentially, to mitochondrial status are those of a “sympathetic nature”, i.e., autonomic nervous system overdrive. This condition can be elicited by chronically over-doing it, where I mean intentionally working or working-out too hard and/or too long.
It can also be triggered by alarm where it is also known as “fight or flight”. Persistent fight/flight demands energy production non-stop, which makes like demands on the body’s microscopic energy apparatus – the mitochondria.
Fight or flight as the reader may know, has evolved to protect the species in times of crisis, be it major or minor. It is not desirable that it be prolonged – yet this is the case for many that suffer from the myriad symptoms that can result. If prolonged, and energy consumption outpaces energy production, it can result in a downward spiral of health and well-being, both mental and physical. Traditional Chinese Medicine, with it’s 5000+ years of history has a proverb for this: “One cannot burn the candle at both ends.”, where the candle has come to be understood as “one’s essence”, or “yin”. Living in a state of chronic sympathetic bias shortens the useful life of our mitochondria, of our cells, of our organs, and of our bodies.
Both heart beat and breathing rates are key indicators of metabolic rate, where both are inextricably linked, breathing rate determining heart rate depending on circumstance and activity. Breathing rate, depth, rhythmicity, & relaxation are the qualities of the breath that determine its affect on autonomic state, where autonomic state is a determinant of metabolic status, sympathetic emphasis increasing metabolic rate and parasympathetic emphasis decreasing it. Paraphrasing, this is where the yogic adage: “One’s life is not determined by one’s years but the number of breaths one takes”, has great relevance.
~
What does this have to do with the body’s electrical relationship to Earth? The answer to this question is that ATP generation and recycling is highly dependent on “electrons”.(9) It is generally assumed that we receive all the electrons we need from respiratory and dietary intake. Yet, the body seems to want to maximize the flow of electrons from Earth “ground” given the chance. Does the Earth “power” life on its surface?
Considering this question, the answer is certainly, “Yes.”, given that all life is nourished in one way or another by the environment that exists at the surface of the planet, taking into account the importance of solar energy. Without solar energy and photosynthesis, there would be no visible life as we know it, but without Earth we would have no place to stand. The question is, “Does planet Earth power life in a literal sense, i.e. electrically?”
The graphs of Figures 7 and 8 demonstrate what happens when the body is engaged in muscular activity, Figure 9 demonstrating rhythmic diaphragm motion of Coherent Breathing, rising from a resting quiescent state over a period of 30 minutes, and Figure 10 demonstrating the muscular actions of the spontaneous kriya, which are “shaking”, arrhythmic movements, during which time various muscles throughout the body are flexing and relaxing simultaneously, the period of the kriya proper lasting about 30 seconds.
In the Coherent Breathing case, current climbs steeply and then gradually over the entirety of the 30 minute period. In the case of the kriya, current snaps abruptly, very much like the rising and falling edges of a square wave, current roughly doubling at its peak. In retrospect, what I have failed to demonstrate in either of these cases is the quiescent current, absent of either deep rhythmic breathing or kriya.
There are two fundamental questions that require answering:
1) Is this current from Earth to the body a window into the mitochondrial “electron transport chain” and mitochondrial status in general?
2) Does the autonomic status of the body shift immediately toward parasympathetic emphasis when grounded? If yes, why?
The patent abstract of 2005 asserts that it does. Furthermore, it asserts that grounding should be applied immediately in urgent medical care situations, specifically for the purpose of mitigating trauma.

Another examination is in order, this being “quiescent” subtle breathing followed by Coherent Breathing, followed by exercise that gradually ramps up in intensity. Does electron flow from the Earth to the body increase during muscular exertion and its concomitant demand for bodily energy production?
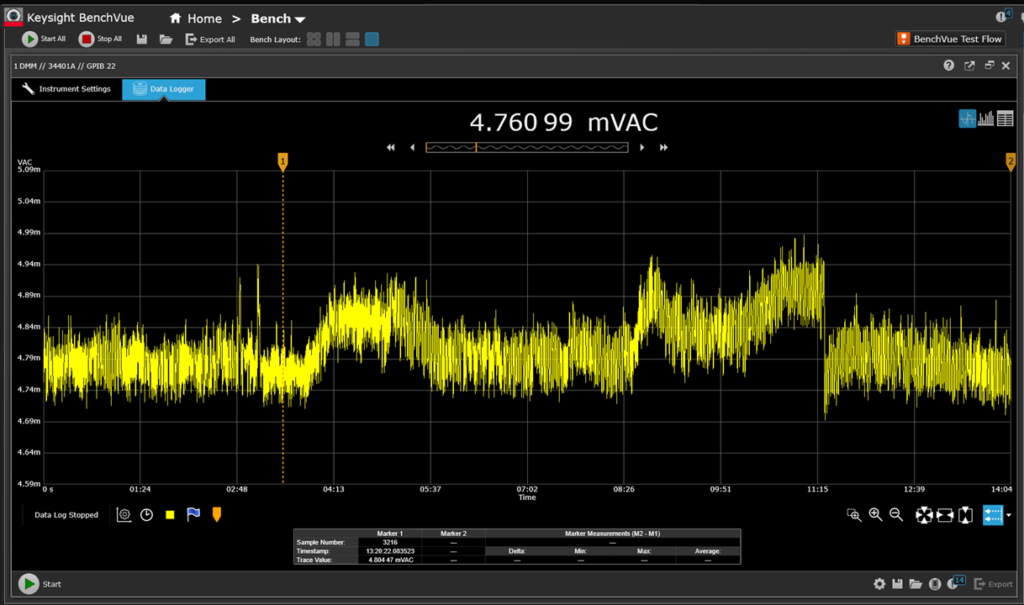
Figure 10: 5 distinct periods of activity with voltage measured across R1 (Instrument: Agilent 34401A)

Figure 11: Starting vales with different instrument: Agilent 34401A
Figure 10 is a new measurement employing an Agilent 34401A meter measuring the voltage across resistor R1 (200 ohms). The presentation and logging is Keysight (previously Agilent) BenchVue. The period of ~14 minutes is divided into 5 intervals. Sampling is occurring at the rate of 16 samples per second.
Note that there is a significant measurement difference between the 34401A and the 3458A instrument used in the initial measurements of early 2000s. The 3458A was very strictly measuring the AC signal only, yielding a starting voltage to ground of .040 VAC. As a $100,000 instrument at the time, the 3458A had the capability to strictly measure the AC component alone, which is where the initial value of .040 VAC to ground came from as well as the table of Figure 5. The 34401 is yielding a “True RMS” measurement of 0.20 VAC between the human body and ground – the instrument being between the two.
As mentioned previously, a best way to validate that this electron flow from Earth to the human body (via the skin) as a function of mitochondrial respiratory chain demand is to test to see if current increases when the body is: a) breathing, and b) working. Figure 10 above is this experiment. The arrangement of the apparatus is the same as that described in Figure 2, and the results previously depicted in Figure 7, i.e. voltage is being measured across a 200 Ohm resistor and recorded.
The 14 minute period is divided into 5 segments:
1) Supine position on the mat breathing very lightly. This period lasts 3 minutes.
2) Supine position on the mat employing Coherent Breathing. This period lasts 3 minutes.
3) Supine position on the mat, performing Nabhi Kriya with alternating leg lifts to vertical position, inhaling while raising and exhaling while lowering, rapid succession. Fatigue building rapidly during the last 30 seconds or so.
4) Supine position on the mat, performing Nabhi Kriya with double leg lifts to vertical position, hands raising and lowering 12 pound weight simultaneously, inhaling while raising and exhaling while lowering, rapid succession. Performed until failure.
5) Supine position on the mat, relaxing and recovering from work.
Current calculated by dividing the voltage by the value of R1, 200 ohms, is presented in Figure 11. Voltage is being sampled 16 times per second, consequently, the current values directly follow voltage values. Here, the particular interest is in the value of the current.
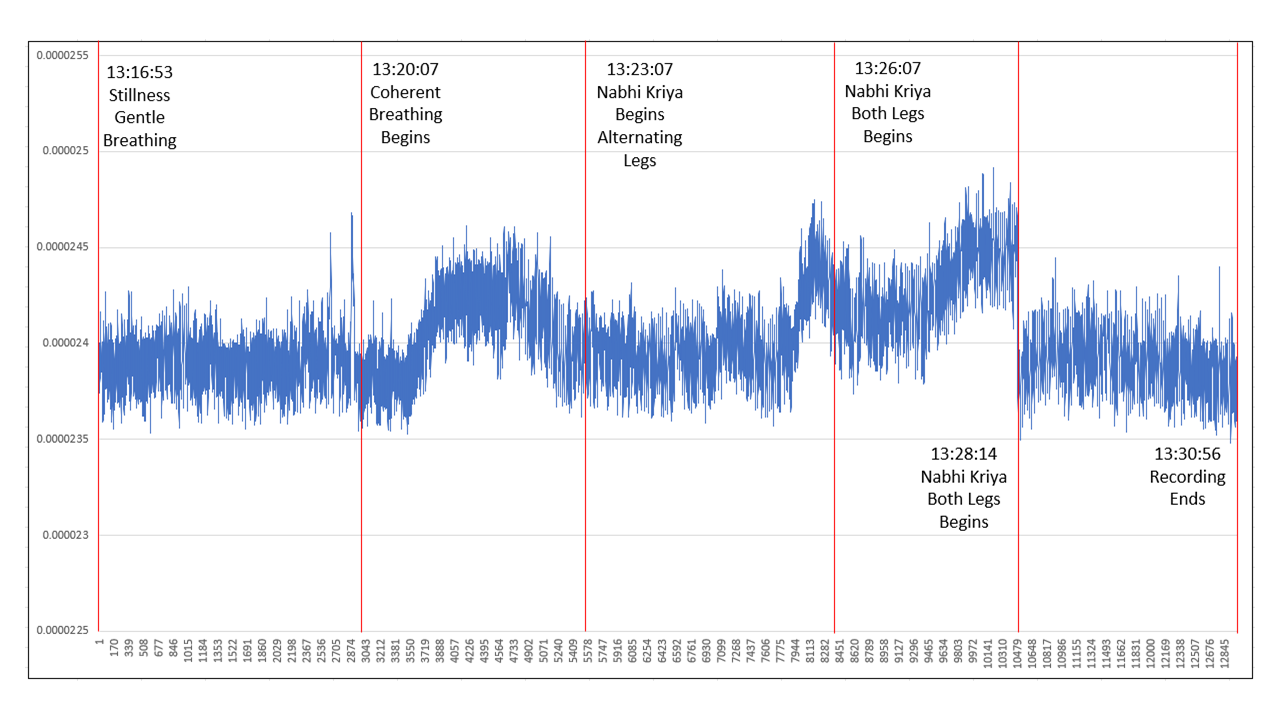
Figure 12: Current as a function of voltage recorded across R1 during 5 distinct periods
The graph is best dissected via a table that presents the averaged values of voltage and current during each period, where “electrons per second” is introduced.
For the benefit of visual comparison, the table is presented first, before the discussion of electrons per second.
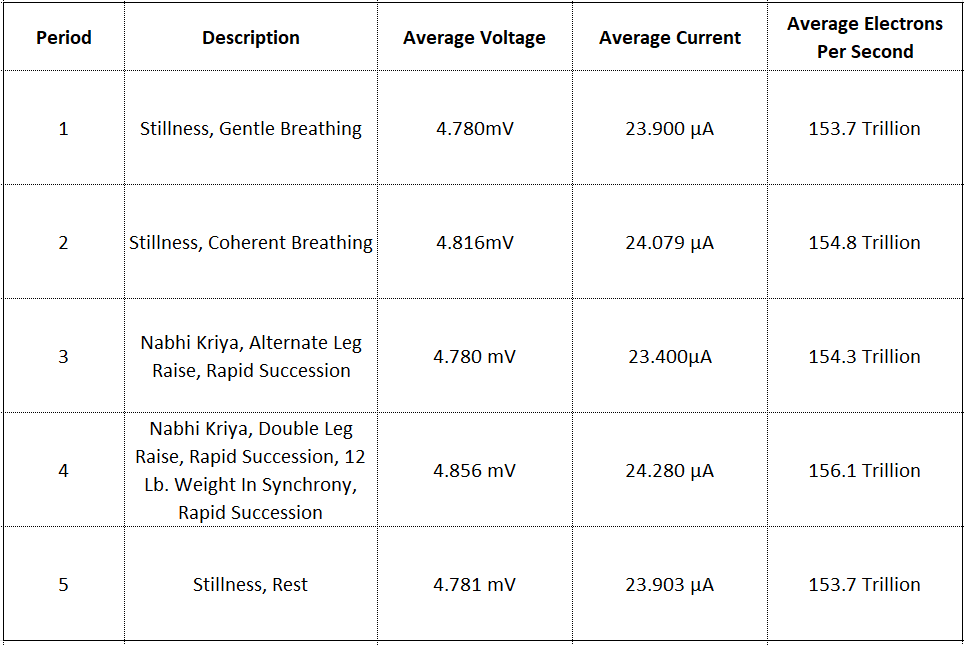
Figure 13: Average current demand during periods of stillness, Coherent Breathing, work, and rest.
By comparing visually, one can see a few things clearly:
Period 1: Current is “nominal” for the first 3 minutes, where electron flow is estimated to average 153.7 trillion electrons per second. A central question of this article is why this current flow exists, where the present hypothesis is that electrons from Earth are easier to obtain via 200 ohms than electrons already in the body, for example circulating in the bloodstream.
Period 2: After a few momentary peaks current trends slightly downward for 10s of seconds after which it begins climbing quickly and continues climbing, after which there appears to be an abrupt adaptation and current begins trending downward to an average that is slightly higher than that of period #1.
While Coherent Breathing continues for the entire 3 minute period, it is as if increased current demand is met and tapers off. We can interpret lower current demand as a consequence of lower voltage, i.e. voltage becomes less positive after a period of roughly 90 seconds of Coherent Breathing. There is a very clear demarcation that occurs at the last 3rd of the Coherent Breathing period like something in the body makes a significant adjustment. (Note that Coherent Breathing is proceeding in synchrony with Vocal Instructive Sequence for this entire period, without excessive motion, strain, or discomfort of any kind.) Electrons per second average 154.8 trillion, the 2nd highest average during the experiment. We will have a closer look at this “shift” in a moment.
Period 3: Current remains at roughly the same level as the end of period 2 until fatigue begins to set in and the burn of lactic acid can be felt, where during the last minute or so current begins peaking, but then tapers off as did the period of Coherent Breathing. Electrons per second during the period average 154.3 trillion, lower than the period of Coherent Breathing, yet the externally visible physical activity level and breathing rate was much higher.
Period 4: After a period of seconds, period 4 activity of dual leg lifts accompanied by diaphragm action and the hands raising a 12 pound weight, all in synchrony begins. Here, current begins climbing again and peaks out for the experiment, the peaking occurring at “failure”. Average electrons per second are the highest for the experiment at 156.1 trillion.
Period 5: Period 5 is simply rest and recovery. Values return to nominal, averaging exactly the same as period 1, electrons per second at 153.7 trillion.
The point of this is to demonstrate that increased work initiates current flow between Earth and the body. With this in mind, we see that there are successive peaks from left to right, the Coherent Breathing peak, the Nabhi Kriya alternating leg peak, and the Nabhi Kriya double leg peak.
A couple of additional interesting observations…
During Nabhi Kriya, while one can control the rate of leg rise and fall, it is generally done at a demanding yet comfortable pace, as one is inhaling during rise and exhaling during fall. The point is that breathing is relatively rapid. Nonetheless, with relatively rapid breathing and exercise, we don’t see much demand for current until the lactic acid burn point is being reached. Why not? While breathing is rapid and movement is rapid, “work” must not be sufficient to drive current demand up until near the end, when exhaustion is approaching.
Period 4 pushes the current to its highest point of the experiment, current climbing from the beginning. Unlike single leg, double leg combined with lifting of the weight with the hands does appear to be enough work to result in demand for additional current. Movement stops out of necessity at the point of “failure”. At this moment, the current drops to its lowest point during the experiment and then rebounds and levels out over period 5, approximating the current of Period 1.
~
In electrical speak, there are two definitions of “current”. First, there is “electron current”, which is electrons moving along a conductor flowing from negative to positive, as electrons themselves are negatively charged, and “conventional current”, the origin of the term being the understanding that “charge” flows from positive to negative and electron flow results therefrom. No matter which term we choose to use, when the body which is “positive” is connected to Earth which is “negative”, electrons will flow from Earth to the body. This is the phenomenon being observed here.
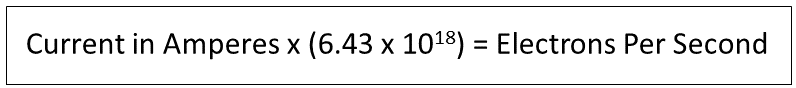
Employing the conversion factor to convert current in Amperes to electrons per second, discovered by American physicist Robert Millikan, for which he earned the Nobel Prize in 1909, we can convert these seemingly small biological current flows into electrons per second, the results being presented in the rightmost column of the table of Figure 12. Here we see that in addition to the relatively constant current of ~23.5 microamps, with each bump upward in current, there is a directly related bump upward in electrons per second.
Here I posit that the electron transport chains of the mitochondrial respiratory chains are reflected in this measurement, that increased ATP production as a consequence of increased work demand is requiring increased electrons over and above the nominal constant current flow of ~153 trillion electrons per second.
Electrons intrinsic to the cell that traverse the electron transport chain are those set free by the splitting of hydrogen atoms into their respective positively charged protons and negatively charged electrons at the beginning of the respiratory chain process, the electrons flowing along the chain and serving to initiate the pumping of proton ions across the mitochondrial membrane at each of 3 intersections in the chain, the addition of each hydrogen ion to the inner membrane serving to increase the positive potential relative to the other side of the membrane.
Internal to the respiratory chain, a very complex electrochemical process occurs, one that took researchers many decades to understand. best explained in this video.(9)
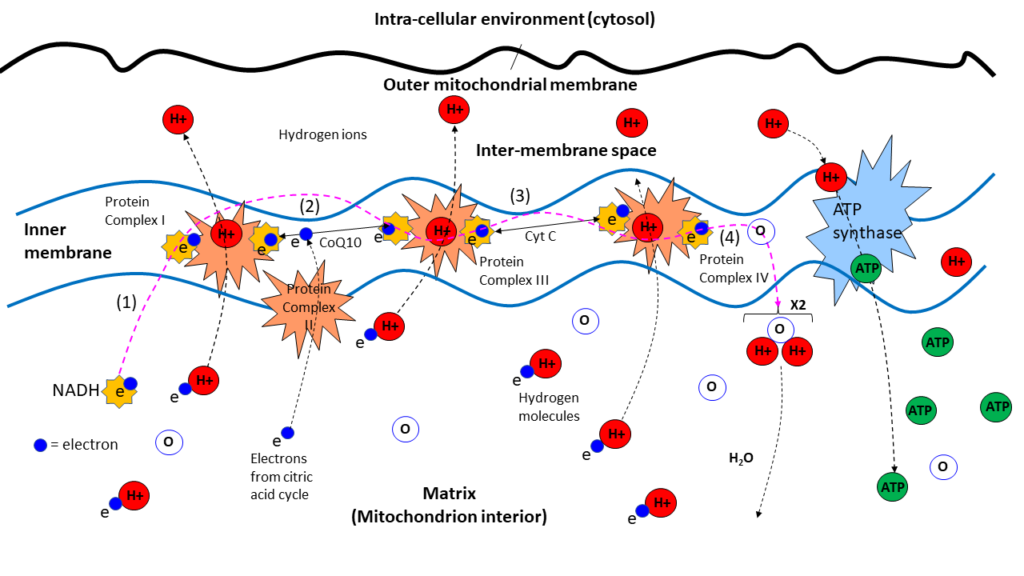
Figure 13: Electron transport chain, a.k.a. cellular respiratory chain.
Referring to Figure 13, it all begins in the cell proper where glucose is broken down into carbon dioxide and negatively charged NADH (nicotinamide adenine dinucleotide), which enters mitochondria protein Complex I where it fuels the beginning of the electron transport chain. The cell excretes the carbon dioxide into the extra-cellular environment where it ultimately finds its way into the venous capillary bloodstream, ending up in the lungs where it exhaled as a biproduct of metabolism.
Upon entering the mitochondria, NADH (negatively charged) reacts with protein complex I to attract a positively charged hydrogen atom lurking in the matrix, the availability of hydrogen atoms being provided via the breakdown of glucose during the citric acid cycle. The hydrogen atom is stripped of its electron and it’s proton ion pumped into the inner membrane space.
A CoQ10 molecule is carried forward in the process by an electron, electrons flowing along the mitochondrial membrane as they would a wire. Each time an electron encounters a protein complex, it attracts a hydrogen atom, strips it of it’s electron, and the protein pumps the resultant hydrogen ion into the inner membrane space where a reservoir of ions build up forming a positive proton gradient between inner and outer membranes. Protein complex II provides additional electrons to the chain directly from the citric acid cycle.
Electrons traverse complex I, III, and IV, pumping hydrogen ions into the inter-membrane space at each stop. Upon entering complex IV electrons combine with oxygen and hydrogen ions to form water, the byproduct of cellular respiration. Of course, oxygen is prevalent in the cell and the mitochondria as a consequence of “breathing”. It is drawn into the body from the photosynthetic environment, “air”, where virtually all of it is produced by plant life, oxygen being the waste product of photosynthesis.
Hydrogen ions in the inter-membrane space then re-enter the matrix via what appears as a crankshaft to which they are attracted due to their charge. The crankshaft catches and carries them though the ATP synthase where they are catalyzed into ATP, chemical energy, which is then released and goes on to “power” the cell and its myriad activities. It is this process that is referred to as “cellular respiration”.
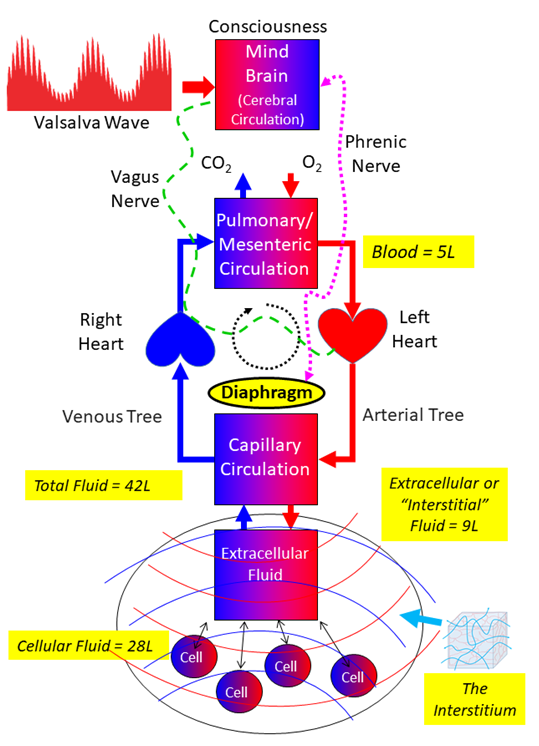
Figure 15: Model Of Circulatory Physiology As Articulated By Stephen Elliott re. Coherent Breathing
Referring to Figure 15 above, inhaling, exhaling, blood moving – in the end is not “respiration”. These are the systemic mechanisms that facilitate the provision of nutrients, gases, waste removal, etc., to aid the production of ATP by the microscopic mitochondria of every cell, which in the end keeps the cells alive, and with living cells, keeps the body alive. If the mitochondria of a cell stop producing ATP, the cell has no source of energy and dies. This is happening to millions of cells in the body every day, dying cells being replaced by new ones via cell division.
“Cellular respiration” is the process internal to cells whereby nutrients provided via the capillary circulation -> interstitial fluid, are consumed by cells, wherein cells break it down and use it to fuel the mitochondria, of which there are 100 per cell (N). The mitochondria use this fuel, to generate a positive polarity between their inner and outer membranes. This is analogous to charging a battery, but in this case the battery, the mitochondria, is nanoscopic, a billion of them fitting inside a grain of sand (N). The reason this can power the workings of a cell and of a mammal, or any other living organism is that there are 1000 respiratory chains per mitochondria, 100 mitochondria per cell, and ~100 trillion cells in the human body. This turns into the number 2 sextillion electrons per second, 2 followed by 21 zeros , the equivalent of 320 amperes of current.
Inside the mitochondria there is an atomic energy production process at work via three fundamental stages: 1) “glycolysis”, 2) the “citric acid cycle” (Krebs cycle), and 3) the “electron transport chain”. What we commonly think of and refer to as “respiration” or “breathing” serves this purpose. Here I explicitly include the role of breathing as a “gas exchange” function, and as a “blood and fluid moving” function, both of which are necessary to the health of the milieu interior, both extra and intra-cellular.
In the cell, glycolysis breaks down glucose into carbon molecules called pyruvates. ATP provides the energy for this process. Note that the root is from the Greek word “pyr”, meaning “fire”. Pyruvates are then converted into acetyl coenzyme A (CoA) which enters the citric acid cycle, where it is converted into carbon dioxide, releasing 2 molecules of CO2, and one each of NADH, FADH2, and GTP.
The CO2 is expired by the cell into the interstitial fluid, on into the venous capillary circulation, and from there to the pulmonary circulation where it is ultimately exhaled during breathing, CO2 being the “exhaust” of combusting carbon molecules in the fire of metabolism.
Recall that it is the negatively charged NADH that enters the mitochondria kicking off the electron transport chain, a series of reactions that release energy at each step in the chain, this energy being used to pump up the positive polarity of the inner mitochondrial membrane, this proton energy being used by ATP synthase to create molecules of ATP.
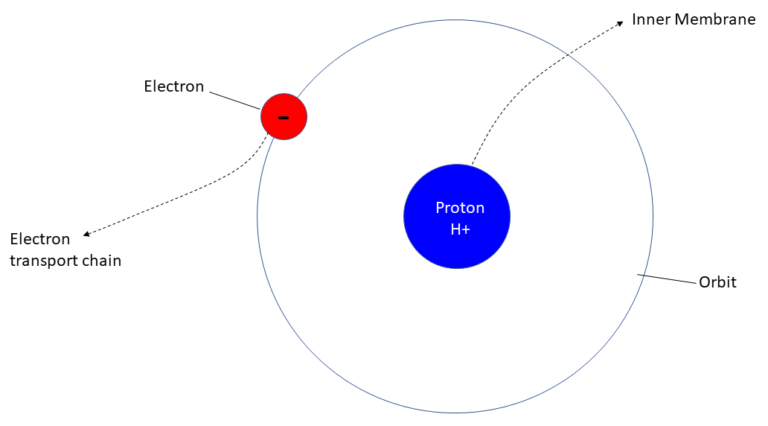
Figure 16: Hydrogen atom and the respective roles of it's proton and electron
Hydrogen has an atomic number of “1”, it has one proton and one electron and is the most abundant element in the universe as we know it, the primary source of hydrogen in this process being water. When hydrogen enters the respiratory chain, it’s single electron and single proton are separated and perform different functions “down the road”, i.e. down the chain. The electrons, wanting to flow from negative to positive are conducted along the membrane, stop by stop, each stop facilitating the stripping of the electron from the hydrogen atom and pumping the hydrogen ion through the wall of the inner membrane, raising it’s electrical potential.
Here we have an example of the most fundamental and pervasive element in the universe as we know it, hydrogen, in the body primarily as H20, water, “splitting”, it’s electron facilitating electrical flow (current), and its proton pushing up the positive voltage gradient across the inner membrane, electrons on one side and protons on the other. The hydrogen that the mitochondria employ is derived from glucose, but the hydrogen in glucose is of water. While the most abundant element on Earth, the gaseous atmosphere of Earth consists of only .00005% hydrogen, so the body’s overwhelmingly primary source of hydrogen is H20.
It is estimated that the human body consists of 2 x 10^25 molecules and that ~99% of these molecules are water. Consequently, ~2/3 of the body is hydrogen and ~1/3 is oxygen. So, we can see that both hydrogen and oxygen molecules are plentiful. Because water approximates 99% of the body, we can also easily calculate the number of electrons that exist in the milieu interior. Needless to say, it is many…
The energy required to “split” a hydrogen molecule is ~13.6 eV (electron-volts). As evolutionary fortune would have it the energy stored in one molecule of ATP is ~7.3 eV, such that 2 molecules of ATP is enough to do the job. Here is a closed loop, ATP powering the splitting of hydrogen, and hydrogen powering the generation of ATP. Again, a way to think of the process is as charging a biological battery, capacitor, or better yet, a fuel cell, where a reservoir of energy is accumulated for purposes of generating ATP, each molecule of ATP being an envelope of chemical energy that the cell can employ in real time to serve its myriad purposes – including splitting hydrogen atoms.
At the end of an electron transport chain exists an oxygen molecule, which accepts an electron and picks up hydrogen molecules, the result being water. Four electrons are required to reduce each molecule of oxygen to H2O. In total, cellular respiration requires six oxygen molecules to break down one glucose molecule into six CO2 molecules and six H2O molecules, the result being ATP and heat.(N) During cellular respiration, one glucose molecule results in 38 molecules of ATP. (N)
This is going on in every cell all the time, but varies in ATP output based on demand. Does this positive proton gradient that exists in every mitochondria in every cell explain the +0.20 voltage presented by the body relative to Earth ground, the starting point of this article? It seems that the product, ATP, carries a negative charge. It is when the highly negatively charged phosphate chain of the ATP molecule is broken by pairing with a positive molecule that it’s electrochemical energy is released. This is what occurs when we flex a muscle or make any other energetic demand on the body, be it somatic or autonomic.
~
The table of Figure 13 is useful to understand the average currents during each of the 5 periods, but by definition it averages out the information regarding the peaks, part of the importance of the analysis. To this end, the table of Figure 17 below makes clear the peak values.
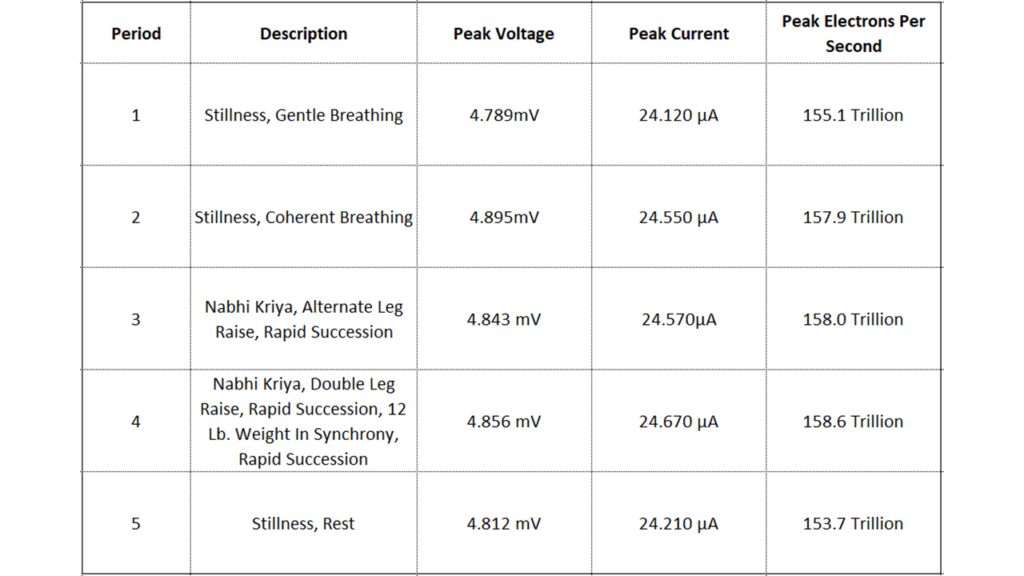
Figure 17: Peak current demand during periods of stillness, Coherent Breathing, work, and rest.
Here we arrive at the opportunity to look more deeply into the signal. Figure 17 zooms into the first 1324 samples or 83 seconds of the “Stillness, Gentle Breathing period, close to half, the total period being 3 minutes or ~180 seconds in length.
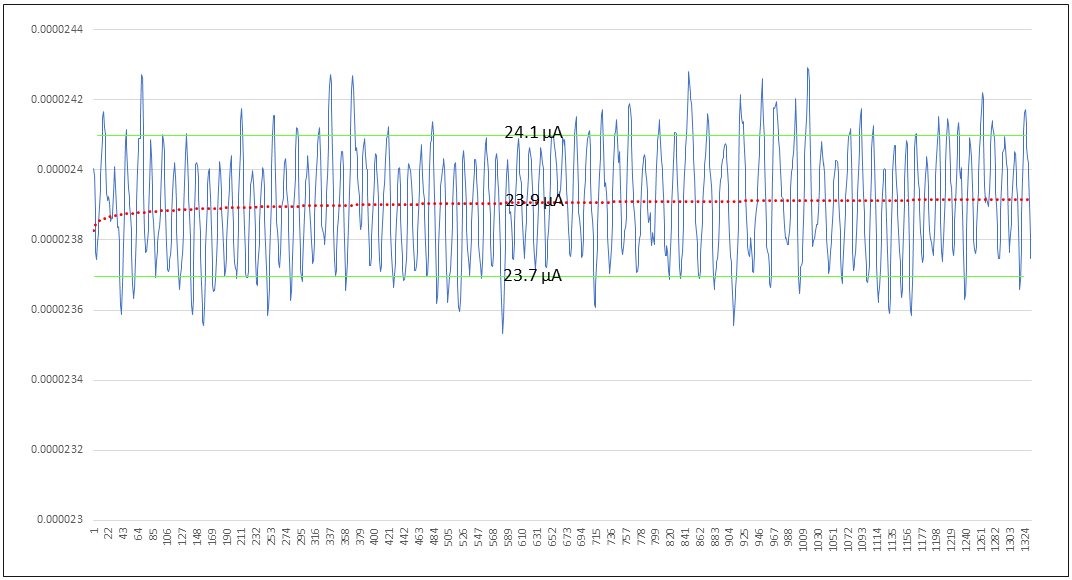
Figure 18: A zoomed-in view of the first 83 out of 180 seconds of Period 1.
What are we looking at? Referring to Figure 15, we can estimate the AC component to be 0.4 microamps, the aspect of the signal between the green lines, and there is a DC component of 23.9 microamps, on which the AC component is riding. The AC component is the relatively small cyclic signal, with commensurately small cyclic electron flow. This AC signal is in fact the heart beat.
The red line is the power trend of the the AC component’s Y axis values, the Y axis is current in Amperes and the X axis is time in “samples per second”, where the sampling frequency of the measurement gear is 16 samples per second. The power trend serves as a best estimate of the DC current across this period. The DC current is the predominant electron flow which is relatively constant at 153.6 trillion electrons per second. Again, why does this current flow exist?
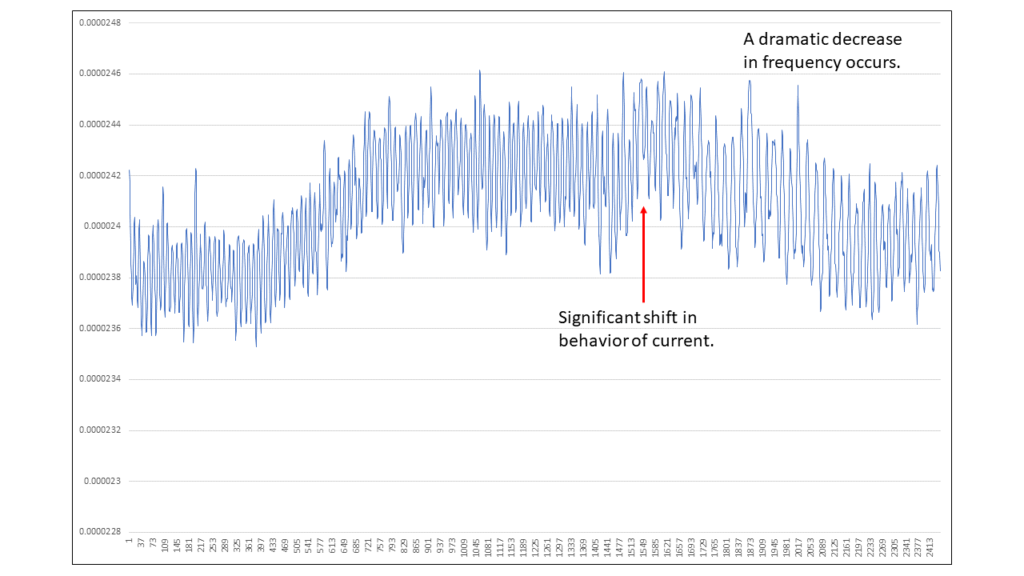
Figure 19: Graph of the entire Coherent Breathing period (only).
Shifting our discussion to the Coherent Breathing period, we see a brief period of quiescent current of ~23.8 microamps and then a rise to ~24.4 microamps, where is stays until a sudden shift occurs at the 143 second mark, frequency slowing abruptly followed by downward trending current ending at ~23.9 microamps.
Here increased current is being provided for a period of time, specifically the period where the heart beat rate is relatively fast, 75 beats per minute, and then it is reduced, specifically when the heart beat rate slows.
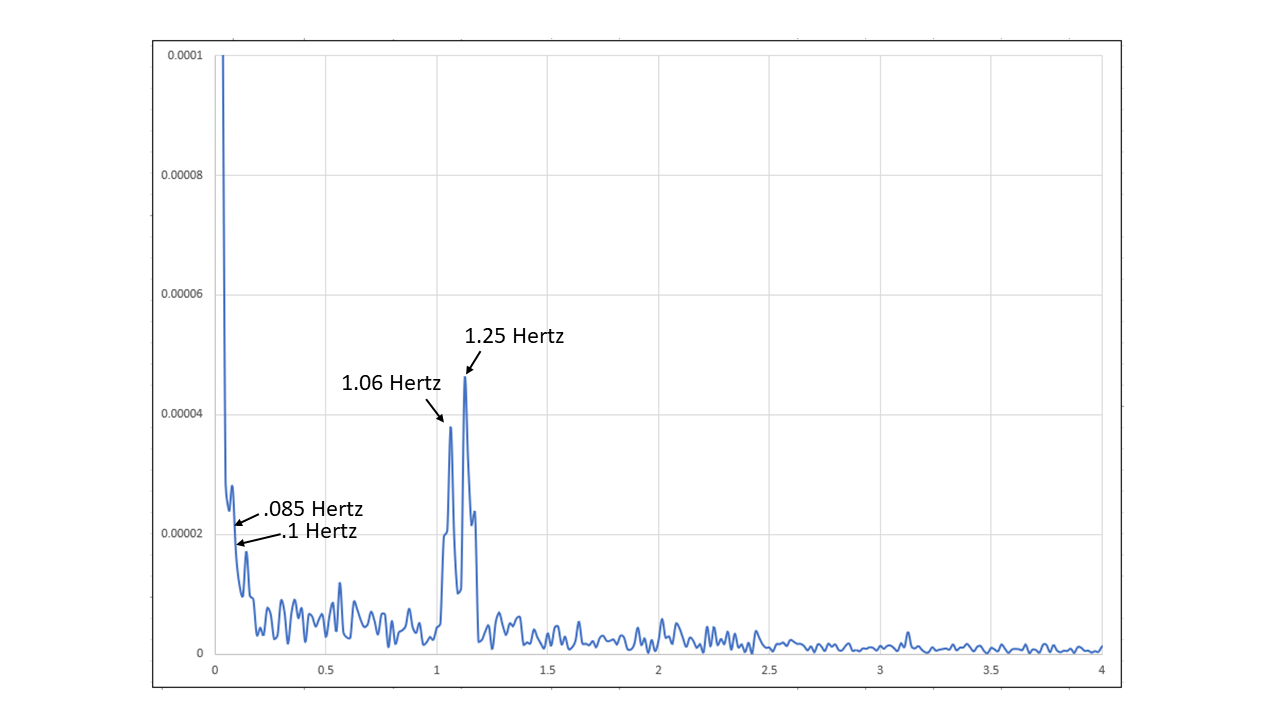
Figure 20: Spectral analysis of the first 143 seconds of Coherent Breathing period. (0-4 Hertz)
The spectral analysis of the first portion of the Coherent Breathing period demonstrates the heart beat at 1.25 Hz., or 75 beats per minute, and also 1.06 Hz. or 64 beats per minute. The Mayer component of 0.1 Hz. is visible as is the breathing frequency of 0.085 Hz.
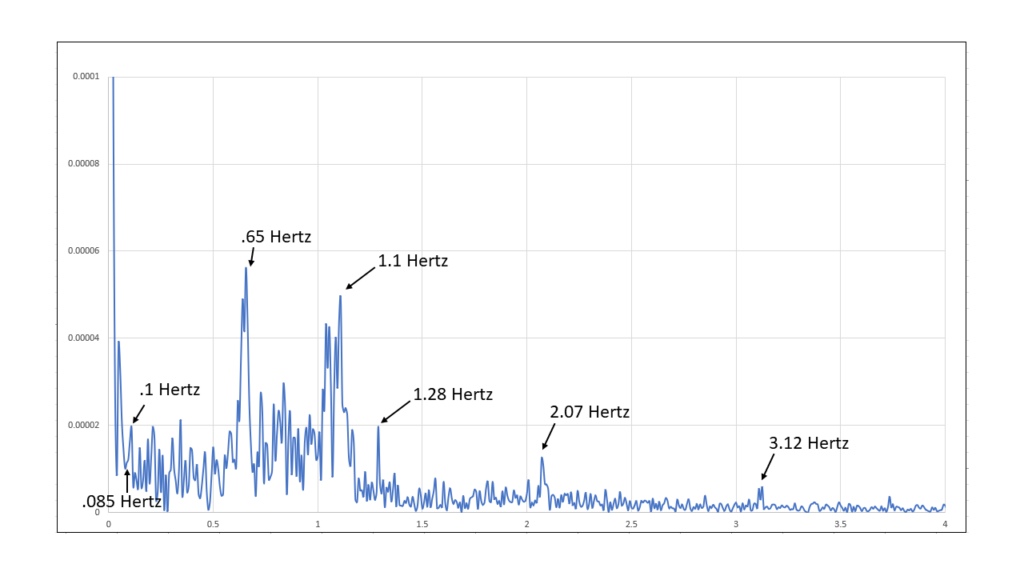
Figure 21: Spectral analysis of the last 37 seconds of Coherent Breathing period. (0-4 Hertz)
143 seconds into the period a significant change occurs, this being the appearance of the 0.65 Hz. component and its harmonics, 0.65 Hertz translates into a heart beat rate of 39 beats per minute, hence, a dramatic slowing down of the heart rate, and with it a trending downward of current and electron flow.
This is the autonomic nervous system shift that we are used to seeing during Coherent Breathing where it is typical that there is a strong shift toward parasympathetic emphasis. A dramatic slowing of the heart rate results in a like slowing of the circulation of blood and fluids to the cells throughout the body, the consequence being a slowing of electron demand. This is a slowing of metabolism that is initiated by breathing deliberately.
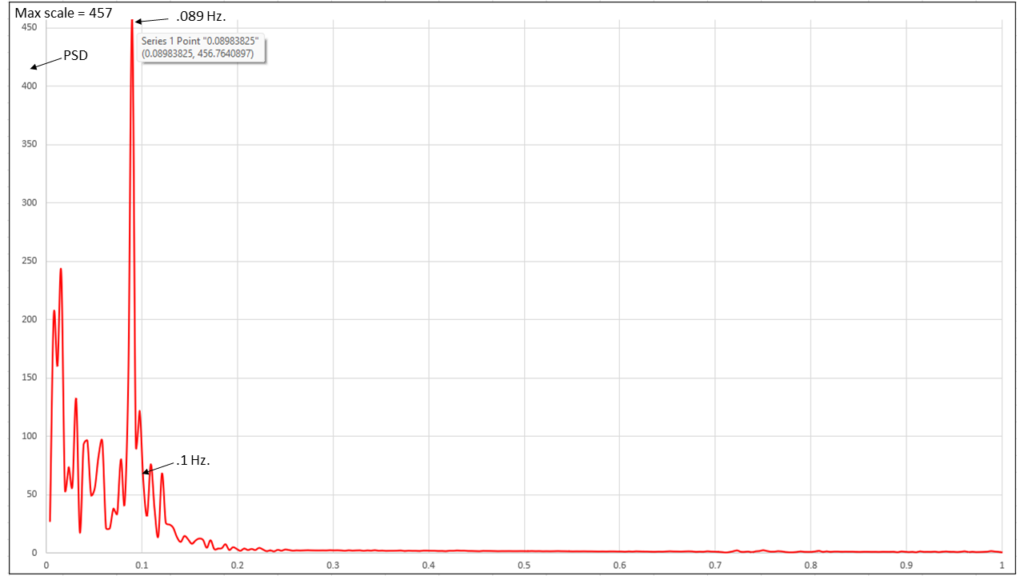
Figure 22: Spectral analysis of Valsalva Wave, 5 min. duration, 1 Hertz. (Instrument: COHERENCE Valsalva Wave Pro)
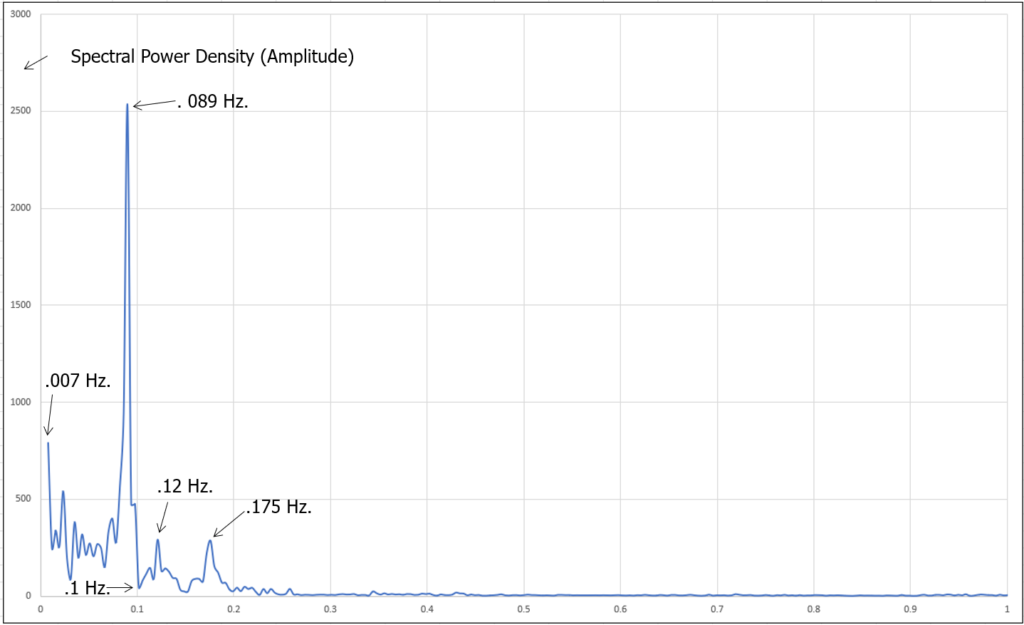
Figure 23: Simultaneous spectral analysis of HRV, 5 min. duration, 1 Hertz. (Instrument: COHERENCE Valsalva Wave Pro)
Figure 21 above shows the quintessential signature of blood activity in the earlobe produced by Coherent Breathing during an electrical assessment. Figure 22, demonstrates the simultaneous HRV spectrum.
This signature accrues from breathing with synchronous rhythm and depth at the nominal frequency of 5 breaths per minute (.085 Hertz), one breath per minute slower than that of the Mayer wave (.1 Hertz), which translates to precisely 6 breaths per minute, the generally accepted frequency of resonance of the adult.
Coherent Breathing is known to produce the Valsalva Wave in the circulatory system, blood volume rising in the venous tree during inhalation and blood volume rising in the arterial tree during exhalation. This wave propagates through the capillary circulation and into the interstitial fluid environment during exhalation, and draws from the interstitial fluid environment during inhalation.
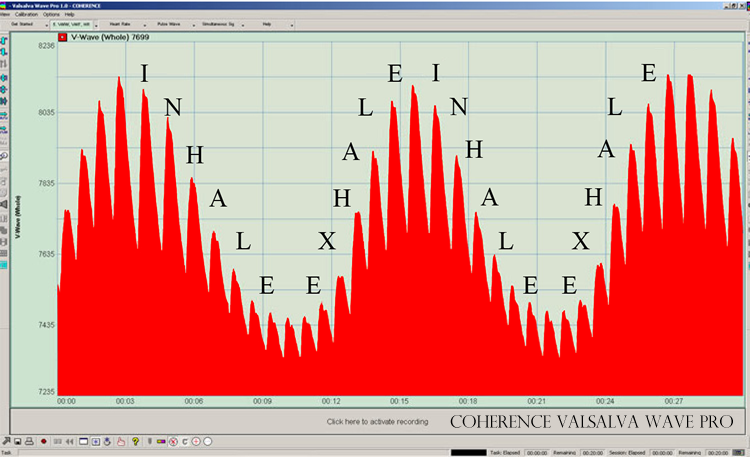
Figure 24: The Valsalva Wave (Instrument: COHERENCE Valsalva Wave Pro)
Inside the cells, the same thing is happening, all a consequence of flows and pressures throughout the body exerted by what this author has termed “the thoracic pump”, down to, and inside cells and their respective organelles (microscopic “organs” that exist inside cells).
This wave is rising and falling at the nominal rate of 5 times per minute. The heart beat is occurring at the nominal rate of 72 beats per minute, however we can see from Figures 18 and 20 that the average heart beat steps downward a notch, and continues slowing until it phase-locks with the breathing frequency, this producing the spectral analyses of Figures 21 and 22.
This is a “resonant” response, the respective systems of the body synchronizing in order to facilitate optimality. Of course the heartbeat riding atop the respiratory wave is creating a pulse with positive pressure (during exhalation) and a pulse with relatively less pressure (during inhalation), internal to the cells and their fluid environments. A consequence of deep inhalation is that it creates a relative vacuum or negative pressure on the vena cava, its venous capillaries, and their respective influence on the extracellular and intracellular fluid environment of the body as a whole.
This is the phenomenon that I posit leads to the slowing of metabolic activity, as can be exhibited by heart beat rate, skin conductivity, brainwave state, and hand temperature, where this has been referred to for almost 20 years as “shifting one’s autonomic state toward parasympathetic emphasis and away from sympathetic emphasis”, ergo, slowing one’s metabolic rate and preventing metabolic run-away.
It is important to note that sympathetic autonomic state is “the human condition”, there being many consequences, a primary one of interest to most is “aging” and age related disease. Aging correlates with metabolic rate and metabolic rate correlates with autonomic state. In previous works I have demonstrated that autonomic state correlates with breathing. Work of others suggests that it also correlates with “grounding”. In this article I will demonstrate that this is so.
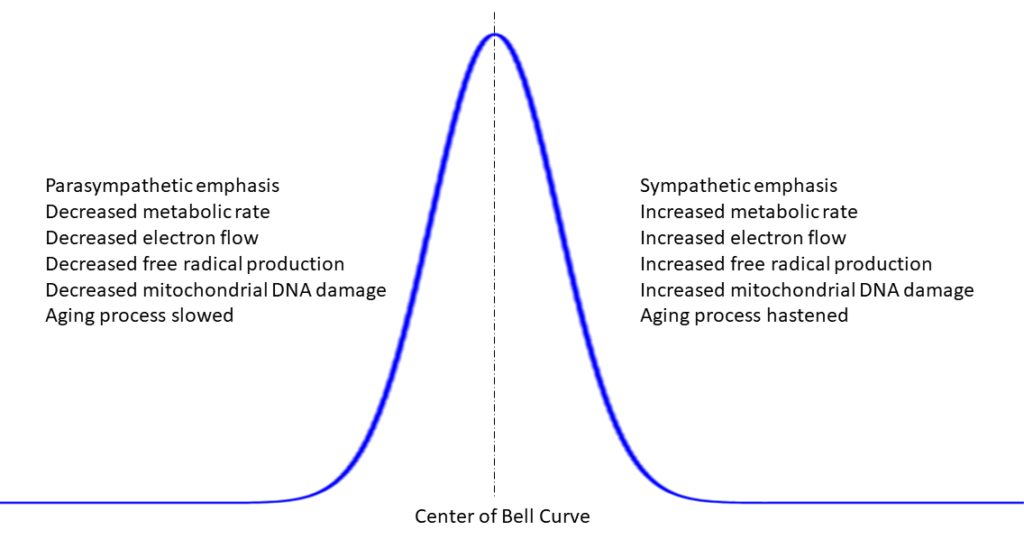
Figure 25: Electron flow in the electron transport chain and the Bell curve
There is a governing mechanism overseeing mitochondrial status and in particular the number of electrons that are present in the respiratory chain relative to energy demand. This mechanism attracts more electrons when they are needed and throttles electrons when they become excessive.
In Power, Sex, Suicide, Mitochondria and the meaning of life (N), Nick Lane describes this action as fitting a Bell curve, where electrons are managed in a closed loop feedback arrangement such that their arrival and utilization stay within a safe zone centered around the peak of the bell, neither too few or too many.
The reason for this is that the electron transport chain works on the basis of reduction/oxidation or “redox”, the term meaning the chemical reaction involving the transfer of electrons between atoms/molecules, either taking or giving an electron at each step in the process, as commonly occurs with the highly reactive oxygen element, i.e. “oxidation”.
When an atom is “reactive” it has a propensity to interact with atoms around it, either giving or taking an electron, changing their current state for better if they need an electron or for the worse if they don’t. This particularly pertains to oxygen atoms/molecules which have a biradical electron configuration, two unpaired electrons, a valence of -2, making it highly available to forming bonds with other atoms and molecules, i.e., highly reactive. Its high reactivity is why oxygen is toxic to aerobic life in high concentration. However, its current concentration of 20.95%, which has remained relatively constant for many thousands of years, spawned the proliferation of fauna as we know it today.
Where did the oxygen come from: The flora that preceded fauna, flora producing all of the available oxygen via photosynthesis.
It is very interesting to note that photosynthesis also works on the basis of splitting hydrogen atoms and proton pumping to create ATP. It is created via the same fundamental process of phosphorylation, but in this case “photophosphorylation”, where the light is the energy that initiates the process, photon energy being converted into and stored as ATP.
~
Reduction then, means the gain of electrons by an atom or molecule and oxidation means the loss of an electron by an atom or molecule, the basis of how the electron transport chain works between complexes.
Referring to Figure 9, the mechanism by which this throttling occurs is signaling between the nucleus of a cell and it’s mitochondria, “mitochondrial-nuclear communication”. The nucleus of the cell detects when free radical formation is excessive as a consequence of excessive electron flow and redox activity and signals the mitochondria to slow down the process of electron flow, i.e., their metabolic rate.
High current results in many redox opportunities and equal opportunities for free radical generation. The term “free radical” may be familiar to most readers and for good reason, the science of the cellular respiration having exposed this potential dagger to health, well-being and longevity 4-5 decades ago. It is currently understood that damage to mitochondrial DNA is the root cause of aging and susceptibility to age related disease.
Early thought on free radical creation attributed it to “oxygen” because the reactivity of oxygen is so well established, hence the flurry of anti-oxidant supplements lining the shelves of one’s local apothecary over the past 50 years. But while oxygen is a contributing factor, it has been proven that oxygen is not the primary cause of mitochondrial DNA damage.
Birds are the primary example of a species with high metabolic rate, high oxygen consumption, yet relatively long lives, some living as long as humans. The mitochondrial DNA of birds does not suffer damage at the rate of other species, e.g. rats, which have high metabolic rates and an average lifespan of about 3 years, their deaths being a consequence of mitochondrial DNA damage and age related disease – the same essential mechanics resulting in human aging and disease.
The foremost reason for this damage is thought to be “excess” electrons piling up near the microscopic location of mitochondrial DNA, electrons possessing the fundamental ability to alter the state of other atoms/molecules, specifically those making up DNA, but also other atom/molecules, whose redox condition can ultimately have a negative impact on the health and integrity of DNA, via a chain of redox events.
~
Energy generation demand may explain the up/down action that we see during Periods 2, 3, and 4 of Figure 11, where it seems that there is a call for electrons and then a throttling back in effect. It is very hard to know because the measurement approach is observing the whole body. At the same time it is useful to know that the energy demands of the whole body might be assessed in this manner.
Numbers are presented below where the conclusion is that the measurement apparatus is capturing a small reflection of what is going on within the cellular respiratory chain, the measurement capturing ~0.0000007% of the total electron flow (Line 52), the remaining 99.999993% of current flow (Line 53) originating within the cells of the body, the electrons being provided by the breakdown of glucose ->NADH, and those arriving directly from the citric acid cycle. Converting the electrons per second down all respiratory chains simultaneously, as if in parallel, yields 320 amperes of current, an astounding number reminding us that pound-for-pound the human body very well may produce more energy than the sun. Let’s leave that for another day…
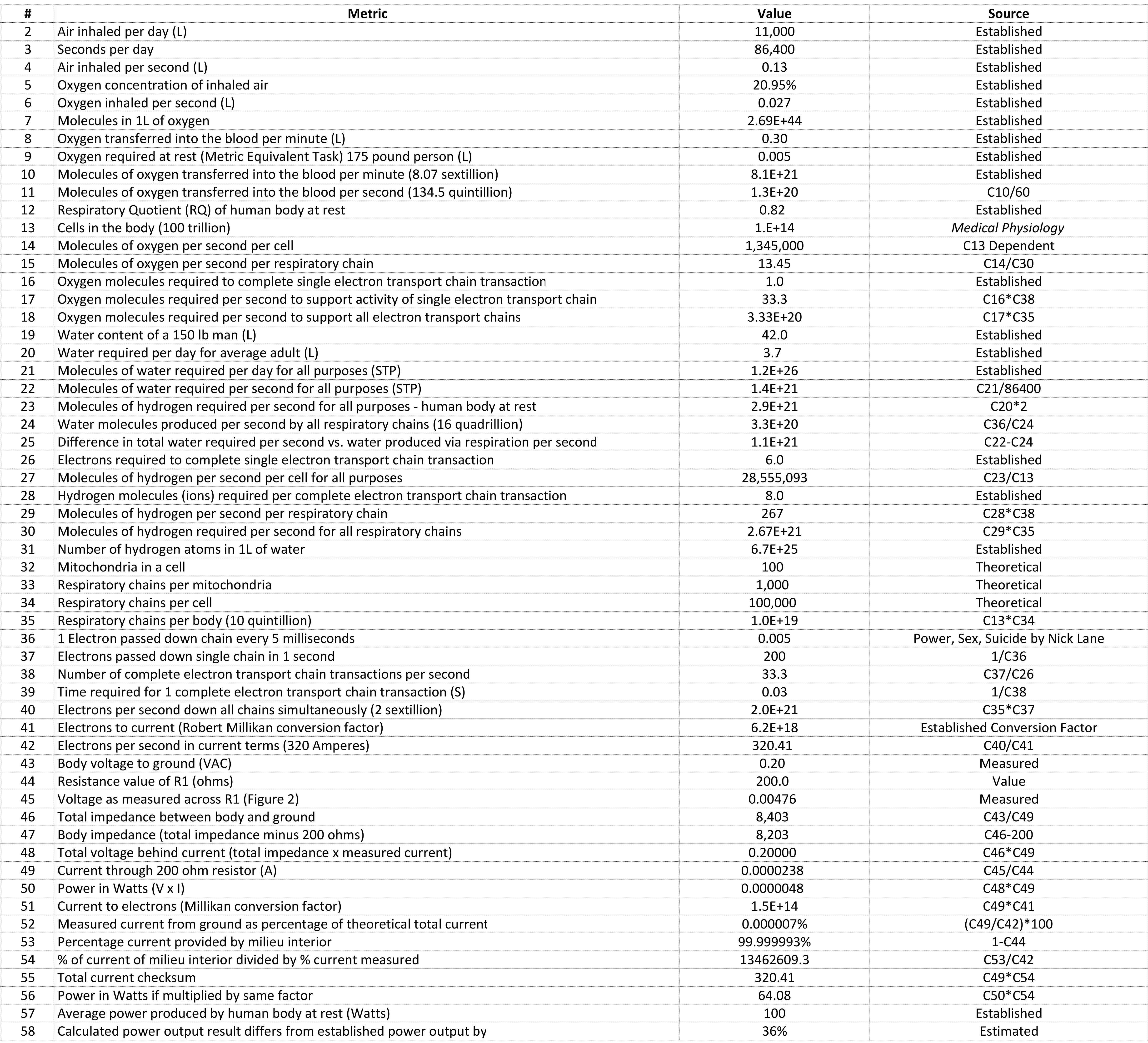
Figure 26: Numbers
A few particularly interesting points about the numbers of Figure 26…
Line 13: Cells in the body = 100 trillion. This estimate comes from Medical Physiology and is the number that I have used consistently for decades. It is acknowledged that there are many estimates floating around, including those as low as 37 trillion. Many of the numbers in the table depend on this value.
Line 16: Oxygen molecules required to complete one electron transport chain transaction = 1.0. This is the oxygen molecule waiting at Complex 4 that ultimately splits and pairs with 2 hydrogen molecules to from 2 water molecules.
Line 17: Oxygen molecules required per second to support activity of single electron transport chain = 33.0.
Line 26: The electrons required to complete a single electron transport chain transaction = 6.0. These electrons equate to current flow.
Line 28: Hydrogen molecules required per complete electron transport chain transaction = 8.0. Compare this to Line 16, oxygen molecules required to complete “a respiratory chain” transaction of “1”.
Line 29: Molecules of hydrogen per second per respiratory chain = 267. This compares to line 17, oxygen molecules required per second equaling 33, 8 times more hydrogen molecules required per second than oxygen molecules. This demonstrates that the body is in effect powered by hydrogen, the source of this hydrogen in the body ultimately being water.
Line 36: The rate at which a single electron is passed along the electron transport chain = 0.005 or 5 milliseconds. (Here I have a reference that says between 5-20 milliseconds, still a work in progress.)
Line 38: The number of complete electron transport chain transactions per second (per chain) = 33.3.
Line 40: Simultaneous electrons per second down all transport chains = 2 sextillion, 2 followed by 21 zeros.
Line 41: Robert Millikan’s conversion factor = 6.2E+18.
Line 42: Theoretical electrons per second flowing in the body in current terms = 320 Amperes.
Line 43: Body voltage to ground = .20 VAC.
Line 44: Resistance value (R1) between body and ground = 200 ohms.
Line 45: Voltage measured across R1 = 0.00476 VAC or 4.76mV.
Line 46: Total impedance between body and ground = 8403 ohms, R1 accounting for 200 ohms.
Line 49: Current through R1 = .0000238 Amperes or 23.8 microamperes.
Line 50: Power in Watts = .0000048 or 4.8 microwatts.
Line 51: Current through R1 converted to electrons per second = 1.5E+14, or 150 trillion.
Line 52: Measured current via R1 as a percentage of total current flowing in the body = .000007%.
Line 53: Percentage of current provided by milieu interior = 99.999993%.
Line 54: Percent of current provided by milieu interior divided by percent of current flowing from ground to body = 13462609.3, demonstrating the tiny fraction of current being provided to the body via R1.
Line 56: Power in Watts (Line 50) if multiplied by same factor = 64.
Line 57: Power in Watts of the human body at rest = 100.
To put this in perspective, a 60 watt light bulb powered by 110 VAC is using 0.54 Amperes. In the case of the human body, it is more like 0.2 volts and 320 Amperes.
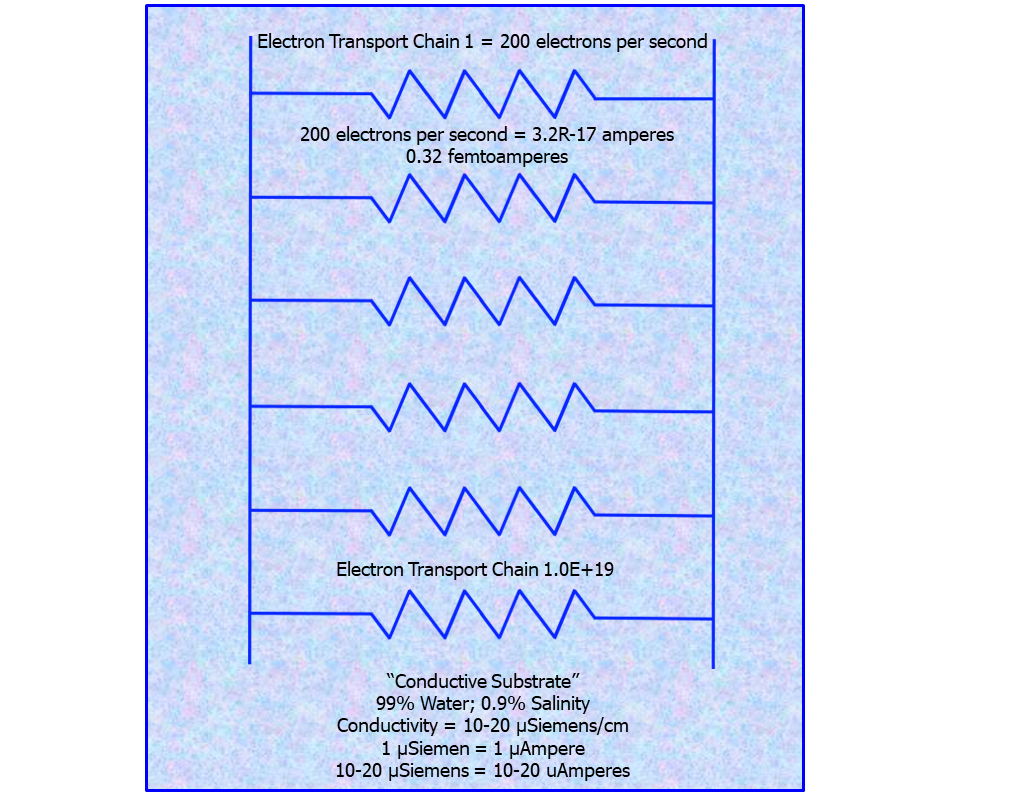
Figure 27: Model of conductive substrate of human body
As we saw in the numbers, the current in all of the cellular respiratory chains adds up to the incredible number of 320 Amperes – this is the body at rest. The body is 99% water. Pure H20 does not conduct electricity and is in fact a good insulator. But the water in the body is laced with sodium, calcium, potassium, magnesium, chloride sulfate, carbonate, and bicarbonate ions.(N) These positively and negatively charged ions make the blood conductive.
If the milieu interior were not conductive, once again, life as we know it would not exist. Of course, this is because energy production relies on conductivity. Without it, there is no electron flow, no positive hydrogen proton gradient, and consequently, no ATP production.
The conductivity of blood is accepted to be between 10-20 µSiemens/ cm, this supporting 10-20 µAmperes of current.(N) Recall that the measurement of Figure 10 averages 24 µAmps. Multiplying 24µA by the value of R1 (200 ohms) yields the average voltages of 4.8 mV, the voltages of Figure 13. It is quite interesting that the apparatus is measuring current that is essentially equivalent to 1 cm of blood conductivity.
~
This has been an interesting foray into the human body and its electrical relationship to ground. This instrumentation allows us a window into the electrical workings of the body and to some degree its metabolic status, e.g. its demand for electrons during rest and activity.
We have also seen that the body is sensitive to impedance to ground, that below 500 ohms current flow between the body and ground is relatively high, but above 500 ohms it drops off rapidly, not linearly. To me this response is telling, and what it is telling us is that the body has a desire for electrons from Earth. Yet, we can see from the numbers that the quantity of electrons the body receives from Earth is extremely small, relative to the total electron activity going on in the body.
So why would the body desire connectedness to Earth, even when the electron exchange is very small (under normal circumstances)?
~
There is an overall problem in the metabolic workings, this being “excess electrons” in the electron transport chains, a.k.a. the respiratory chains. When there are excess electrons, they can pile up and spill out of their “wire” in the vicinity of mitochondrial DNA where they can react with other atoms/molecules in a redox manner. This “wire”
is an imaginary term to describe the path that electrons in the cellular respiratory chain follow. It is formed from a series of “electron carriers” which catalyze the transfer of electrons from reduced coenzymes to oxygen, thereby forming water.(N) The electron carriers in the respiratory chain are arranged in order of increasing negativity, thereby amplifying their negativity such that they have the charge to facilitate the split of the oxygen molecule waiting at complex 4.(N)
The path is not that of an insulated wire, but of an organic pathway along which electrons skip. Consequently, if the arrival of electrons exceeds their utility in the moment, they can spill out of the pathway into the mitochondrial interior. The problem is that they present the risk of damage via their redox potential, especially as it relates to mitochondrial DNA. This damage adds up over a lifetime resulting in “aging” of the mitochondrial DNA, and hence, aging of the organism as a whole.
For many decades the theory of mitochondrial aging (N) posited that free radicals, as a consequence of excess electrons, are the root cause of aging, and that antioxidants would combat their effect.
Nick Lane argues that this has been proven to be incorrect, that given a healthy diet, cells have the natural ability to create antioxidants to combat oxidative activity, and that there is no proof that antioxidant supplementation wards off aging, that the only thing that has been found to slow the aging process across species is dietary restriction ― eat less and live longer.
It is understandable why this works. If we eat less, we can perform less work, due to the fact that the body does not have the fuel to perform work. Hence, the mitochondria are not called upon to produce ATP, extending their useful life cycle. (There is a cautionary point to be made here, this being that if we eat less yet work more, we drive the body into a state of deficiency which ultimately results in sympathetic emphasis and mitochondrial overdrive. This is a reason why it is important not to overdo it while fasting.)
Yet regarding aging, over time the battle is lost.
So, long healthy life all boils down to optimization of cellular respiration and the behavior of the electron transport chains of the mitochondria, where a quest must surely be optimization of electron flow, staying within the confines of the center of the Bell curve – “the middle way”.
~
So, what are the mechanisms at work that result in deviation and what can we do about them? Here I agree with Clinton Ober that a primary answer to this question is “noise”, but I will offer a different explanation.
The body is highly electric and cells are functioning on extremely small potentials, potentials resulting from internal electro-chemistry, and potentials resulting from environmental exposure.
Current flow creates an electromagnetic field that rises and falls with the flow of every electron. The rise and fall of trillions of simultaneous electromagnetic fields generates noise that can be induced in what I will refer to collectively as the conductive substrate of the body.
The noise generated by one electron transport chain can interfere with processes in neighboring electron transport chains – resulting in suboptimal transport chain regulation, i.e. operation outside the thresholds of safety on either side of the peak of the Bell curve, fewer electrons than are needed and more electrons than are needed at any millisecond in time.
This exact problem exists in large scale digital systems, where many thousands of devices are switching on and off, often all synchronized to one master clock signal. With every “on” and “off” an impulse is generated that is reflected in the signal and can be observed on the rising and falling edges of square waves as well as on the “power rails”, where the sum of impulses is visible.
The phenomenon exists at the sub-system level, circuit board level, and is especially critical at the chip level, i.e., inside high density low-voltage integrated circuits, where this impulse noise and associated transmission line effect had to be tamed in order to increase density and speed – where the result of this noise was digital errors, from an information processing perspective, “information errors”. An answer to hardware induced information errors is “error detection and correction” mechanisms, e.g. parity bit checking, check summing, cyclic redundancy checking, and ever more sophisticated approaches and algorithms.
An important principle in telecommunications is that “noise” ultimately limits the information that can be transferred over a channel, where the noisier the channel the lower the information throughput, and the more “processing” of the received signal is required to eliminate errors. Here we are talking about the integrity of any signal, be it analog or digital, but I will focus on the integrity of “1s” and “0s” for now.
Referring back to large digital systems, this phenomenon exists at the sub-system level, circuit board level, and is especially critical at the chip level, i.e., inside high density low-voltage integrated circuits, where this impulse noise and associated transmission line effect had to be tamed in order to increase density and speed – where the result of this noise was digital errors, from an information processing perspective, “information errors”. An answer to hardware induced information errors is “error detection and correction” mechanisms, e.g. parity bit checking, check summing, cyclic redundancy checking, and ever more sophisticated approaches and algorithms.
An important principle in telecommunications is that “noise” ultimately limits the information that can be transferred over a channel, where the noisier the channel the lower the information throughput, and the more “processing” of the received signal is required to eliminate errors. Here we are talking about the integrity of any signal, be it analog or digital, but I will focus on the integrity of “1s” and “0s” for now.
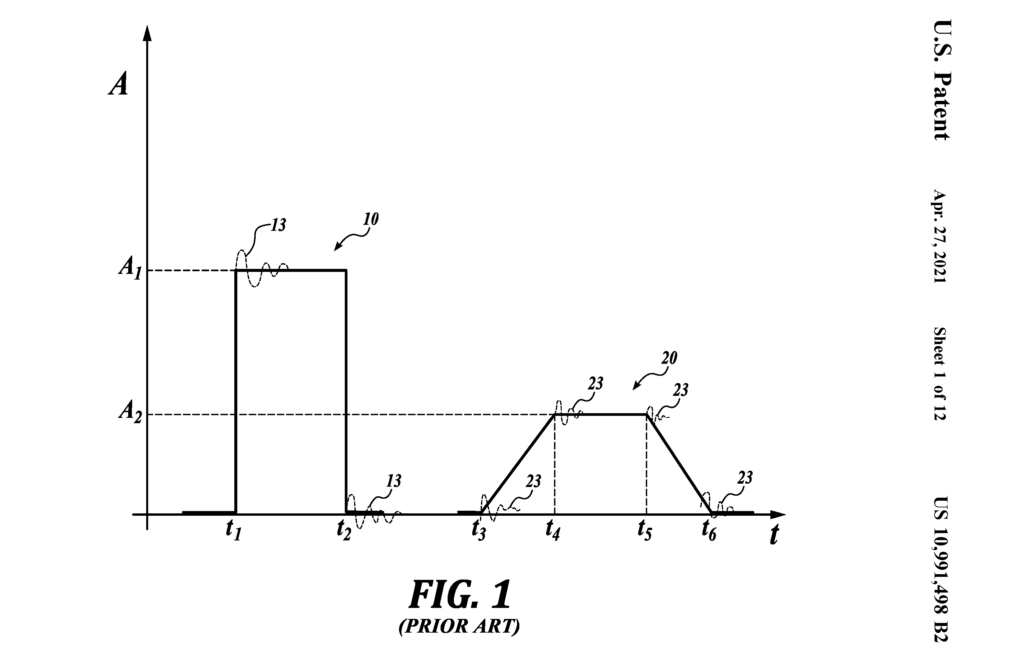
Figure 28: Noise associated with square waves. (US Patent 10,991,498)
Figure 28 demonstrates this phenomenon. The first pulse is a square wave proper. The 2nd pulse is a trapezoidal pulse, a step the industry took to tame impulse noise. It works by slowing the rise and fall of the edges, but still demonstrated impulse noise. It is interesting to note that this “impulse” exists across all forms of flow, i.e., electrons, fluids, air, e.g., electronics, hydraulics, pneumatics. In the last few decades the electronics industry has gone to great lengths to tame this phenomenon because it limits system size, speed, and hence, capacity. This was accomplished primarily via impedance matching.
In 2017, this author, et al, went a step further, positing the “sine pulse”, a pulse that can still have infinitely fast switching speed but switch completely without the generation of impulse. Surprisingly, this invention was still without U.S. Patent Office claim at the time of filing.
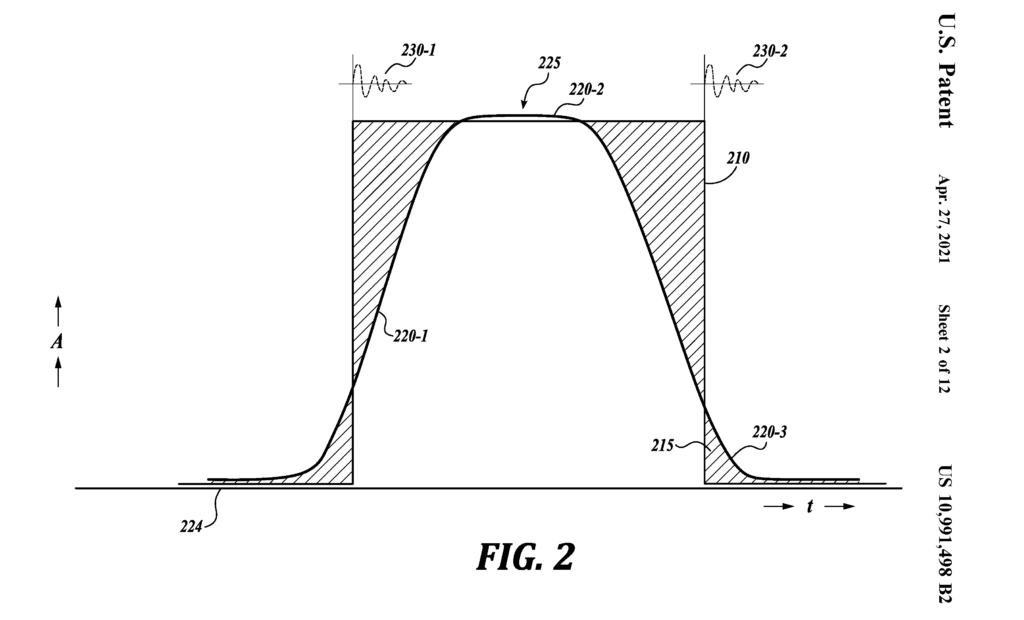
Figure 29: The Sine Pulse (US Patent 10,991,498)
Figure 29 presents Figure 2 of US patent 10,991,498, the “Sine Pulse”. Here the edges are rising and falling sine functions, the sine wave being free of abrupt “on” and “off” states, and therefore free of the infinite odd harmonic noise created by “switching”. In concept, it has an infinite number of applications, but comes with its own complications, this because it has “analog” features.
Readers may be familiar with this author’s fascination with the sine wave as it relates to the internal workings of the human body, where I have referred to it “as the signature of nature”.
~
So, what are the mechanisms at work that result in deviation of current flow in the human body and what can we do about them? Here I agree with Clinton Ober that a primary answer to this question is “noise”, but I will offer a different explanation.
The body is highly electric and cells are functioning on extremely small potentials, potentials resulting from internal electro-chemistry, and potentials resulting from environmental exposure.
Current flow creates an electromagnetic field that rises and falls with the flow of every electron. The rise and fall of trillions of simultaneous electromagnetic fields generates noise that can be induced in what I will refer to collectively as the conductive substrate of the body.
The noise generated by one electron transport chain can interfere with processes in neighboring electron transport chains – resulting in suboptimal transport chain regulation, i.e. operation outside the thresholds of safety on either side of the peak of the Bell curve, fewer electrons than are needed and more electrons than are needed at any millisecond in time.
This exact problem exists in large scale digital systems, where many thousands of devices are switching on and off, often all synchronized to one master clock signal. With every “on” and “off” an impulse is generated that is reflected in the signal and can be observed on the “power rails”.
In the human body, the master clock is the heart beat, however, the master clock of the heart beat is the rate at which the diaphragm moves, which is under control of the phrenic nerve emanating from the brain stem. Here, as conscious beings we have the choice of leaving the diaphragm master clock to run on its own, or aid in its regularity. Of course, this discussion would be incomplete without mention of the Mayer Wave, a pulsatile signal generated at numerous locations in the nervous system that facilitates contractile motion of the arterial tree, propagating the flow of blood (and fluids), every minute of the day, 24 hours a day, 7 days a week. If we were to choose a “master clock” at which the human body operates, evidence suggests that it would be the Mayer Wave function, operating at 0.10 Hertz. Both breathing and the Mayer Wave produce sinusoidal signals in the circulatory system.
This offers clues as to the innate preference that the body has for sinusoidal action of blood and fluid flow.
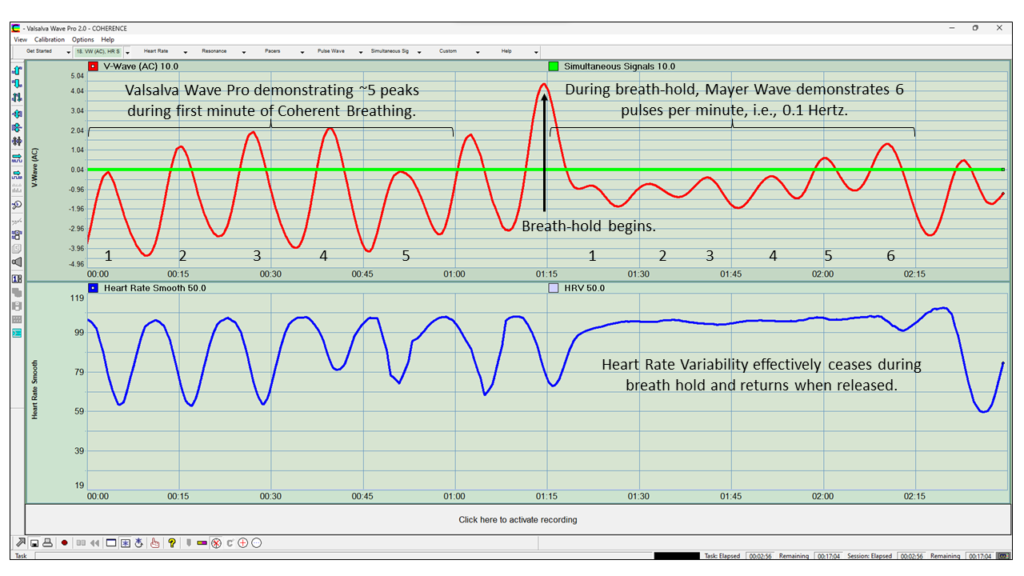
Figure 30: Sinusoidal Pattern Generated By Both Breathing And Mayer Wave
Figure 30 demonstrates that both breathing and the Mayer Wave result in sinusoidal activity of blood flow (red graph). This sinusoidal action resulting in rising and falling pressures and flows across the pervasive capillary membrane, moving both blood and fluids in a “natural” and graceful manner, this wave being experienced by the capillary circulation throughout the body’s ~24,000 miles of microscopic blood vessels, particularly the brain which is held above the chest when in an erect body position. The brain possesses oscillatory functionality that results in electromagnetic emissions that can be observed at the surface of the scalp. Functional brainwave bands consist of frequencies ranging from 1 Hertz to 30 Hertz. Via biofeedback, the amplitude of various frequencies at different locations of the brain can be trained to increase or decrease, thereby affecting a change in an individuals functioning. An example of this is brainwave training for ADD and ADHD.
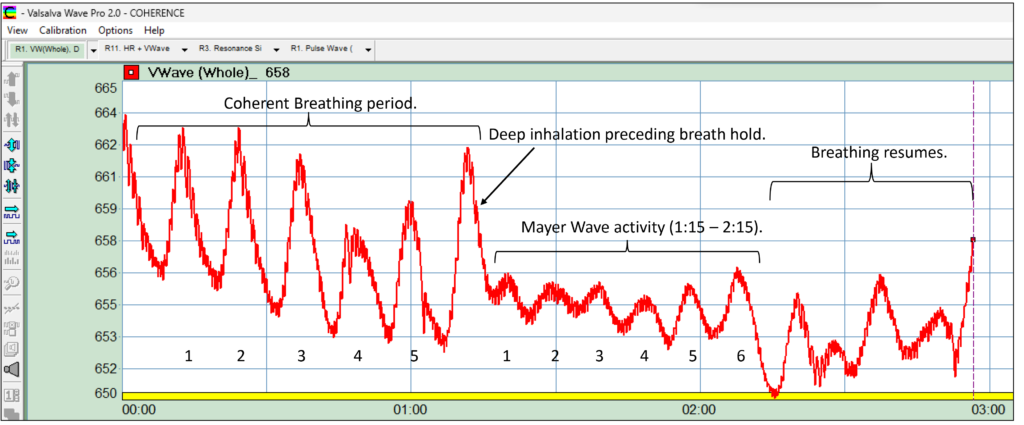
Figure 31: Both Valsalva Wave and Mayer Wave demonstrating sinusoidality
Figure 31 presents the “raw” signal from which both Valsalva Wave and heart beat rate are derived, the heart beat riding atop throughout, the sinusoidal pattern being self-evident.
~
Mainframe computers and digital switching systems of the “digital age”, 1970s to present, were all confronted with the issue of noise, where the more advanced the silicon fabrication, the more challenging the issue. This was all a result of the innovation and electronic adoption of the revolutionary “flip-flop”, a digital element that flips from a “0” state to a “1” state given the state of inputs and a clock signal transition. Given a different state of inputs, it flips back from 0 to 1. The flip-flop is an early ancestor of the mathematical model of computation referred to as the finite state machine. From it, more sophisticated state machines including those of present-day advanced microprocessors could be developed.
The problem introduced by flip-flops is that they and their evolutionary outcomes, i.e. shift-registers, counters, memory chips, arrays, microprocessors, etc., work on the basis of central clock signal that is distributed throughout very large systems, hence we have many millions of flip-flops all switching from 0 to 1 or 1 to 0 on the same edge of the clock signal, i.e., rising edge or falling edge. This synchronous switching of many millions of flip-flops results in what is known in the industry as Synchronous Switching Noise or “SSN”.
SSN was an impediment to the speed at which switches could “switch”, and hence, was an impediment to system size and speed, where the faster the rise and fall of square waves, the more powerful the impulse. The reason for this is that “switches” by definition turn on and off, “connecting” and “disconnecting” such that current will be either allowed to flow at 100% or not to flow, again at 100%. This is in keeping with Newton’s first law, that objects at rest will remain at rest and objects in motion will remain in motion unless acted upon by an external force. In this case, it is electrons that are starting and stopping, and they resist, creating theoretically infinite odd harmonic content, this harmonic content presenting itself as “noise”. A visible example of this is the 110 volt light switch that sometimes sparks when it is flipped, especially when it is flipping from “on” to “off”, where all of a sudden a gap is created and electrons try to bridge the divide.
These obstacles were overcome, leading us to the systems and services on which we rely today, an example being 1Gb/S fiber optic internet service, and for that matter the internet itself, which relies on massive, highly complex hardware/software machines we commonly refer to as routers. These systems could not exist had the “SSN” of the 1980s not been addressed. This being said, as chip switching speeds increase, as they will continue to do into the future – on their own Moore’s Law, management of SSN will continue to be a requirement, necessitating innovation to control SSN in parallel with the innovation that increases switching speed.
Leading technology company’s circuit design CAD applications have this analysis and mitigation capability built-in. By this I mean that every connection on a board or chip is analyzed for signal integrity, transmission line effect, and potential for both emitting and receiving noise – before it is physically cast.
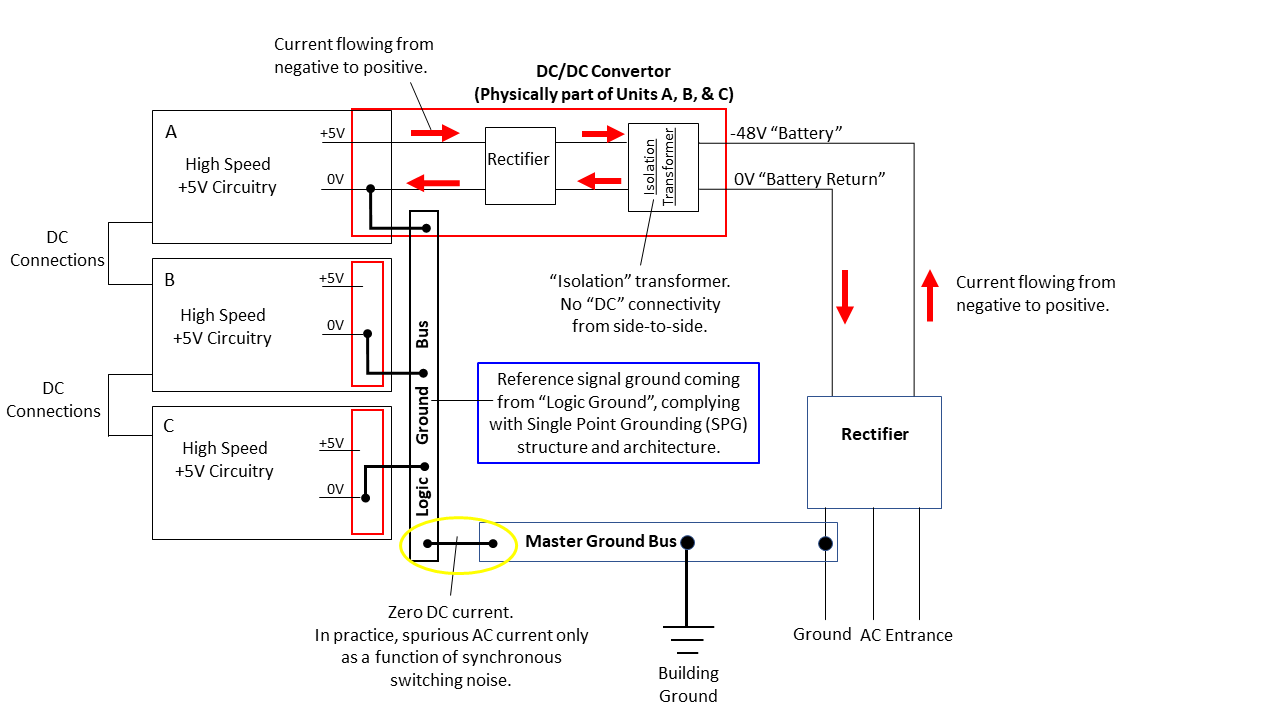
Figure 32: Grounding architecture of high current synchronous digital switching systems
Here I bring your attention to Figure 32, the industry approach to providing digital telephony services with 99.999% availability, a requirement of the US Federal Communications Commission for many decades (sadly, now gone by the wayside due to industry pressure).
The fundamental means of avoiding and mitigating SSN was via a very specific grounding architecture – an electrical grounding design known as “Single Point Grounding – SPG” that would minimize the consequence of synchronous switching noise, and more importantly, prevent oscillating “ground loops”. A requirement of SPG is that ground circuitry branch out like a tree, but with limbs never touching. In practice, this was implemented with multi-layer circuit boards and chips, with complete layers being dedicated to 0-volt power plane and logic ground planes.
Referring to Figure 28, the first thing to note is that the DC-DC Convertor isolates the power input side from the power output side via transformer. Every “subsystem” in a SPG architecture is AC coupled and DC-isolated from every other subsystem, i.e., there is no DC continuity between subsystems – all signals are AC-coupled.
There should be zero DC current flow on the “logic ground bus”, or from it to the “master ground bus”. All of the DC current is sourced from the DC/DC Convertor. The logic ground provides an electrical “reference” to Earth ground, serving to hold the “ground plane” of the system, the 0-volt power rail, at “0” volts.
(Adequate power distribution is also required for this to work, i.e. conductor sizing throughout the system must present zero ohms of resistance to the current flowing from the DC/DC Convertor zero-volt output to devices and from devices to the +5V supply – conductor sizing throughout the system must present zero ohms. These are necessary design considerations for a system to be free of noise.)
Assuming that 0-volts and +5 volts is equally distributed to A, B, & C, if we remove the logic ground connections from A, B, & C, they continue to function but are now “floating” relative to Earth ground, this making them subject to both receiving and emitting electromagnetic interference, this EMI having the potential to effect neighboring circuitry, the higher the frequency and switching speeds, the greater the potential for emissions.
If we were to remove the Logic Ground Bus from circuit “B” alone, its DC connections to circuits A and C (which are technically not allowed to exist – but I add them to help illustrate the point) would become problematic, because B is floating and A and C are not.

Figure 29: Human electrical system
Here I want to draw the parallel between “digital systems” and the human body/mind. The problems I describe above are those of systems operating at the nominal voltage of +5 and single signal currents in the 1 milliamp range.
Using numbers from the table of Figure 26, the human body is operating on voltage(s) of .2 or less and currents in the femtopampere range, dividing Line 37 of Figure 26, 200 electrons per second, by Millikan’s conversion factor yields 3.2E-17 or 0.32 femtoamperes, extremely minute current flows and what I will risk to call “signals”, in that they have an electrical “signature”.
Relative to electrical errors related to noise, what does it take to flip a “1” into a “0” in the human brain? Putting it in language terms, a “yes” into a “no”; in terms of thought, “clarity” vs. “confusion”; in terms of emotion, “harmony” vs. “angst” – parasympathetic vs. sympathetic….?
To the degree that connectedness with Earth can aid humanity in these matters…
~
Albert Szent-Gyorgyi, a Hungarian physiologist who won the 1937 Nobel Prize for the discover of vitamin C is credited with the quote: “Life” is nothing but an electron looking for a place to rest,”(N), the point being that life is fundamentally electrical and rests on the flow of electrical current, i.e. electrons.
The topic of “cellular respiration” and the quest to understand how it worked was a highly sought after holy grail of biochemistry for over a century. Because the interior of the eukaryotic cell contains many microscopic structures, and microscopes themselves where undergoing technological evolution to ever greater magnification, and because of the puzzling biological process that was taking place in the mitochondria, it took many decades of scientific pursuit and argumentation to understand and finally confirm, the advent of the electron microscope in 1939 aiding in this process.
ATP, adenosine triphosphate, had been discovered as the chemical energy being produced by the mitochondria in 1929, ATP being the source of energy for all other cellular processes, but the very means by which it was being produced was the incredible mystery.
Editing....
Footnotes & References
Undergoing editing…
Appendix
Undergoing editing…